

Evolutionary restoration of fertility in an interspecies hybrid yeast, by whole-genome duplication after a failed mating-type switch
- PMID: 28510588
- PMCID: PMC5433688
- DOI: 10.1371/journal.pbio.2002128
Evolutionary restoration of fertility in an interspecies hybrid yeast, by whole-genome duplication after a failed mating-type switch
Abstract
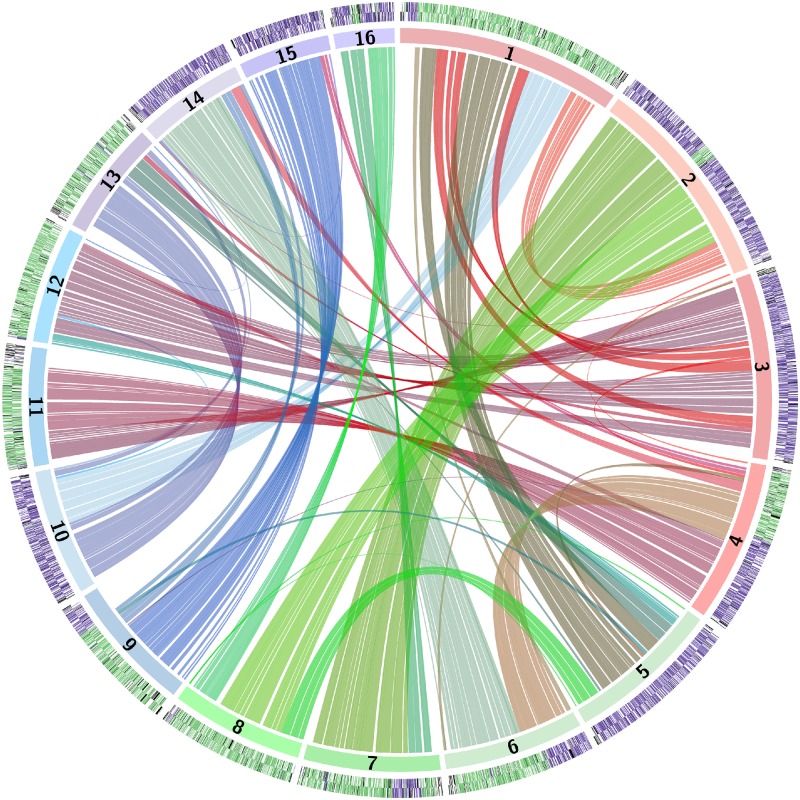
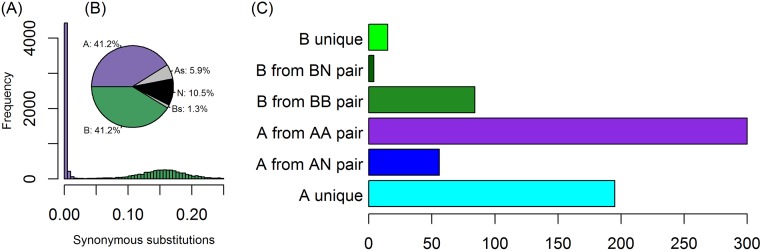
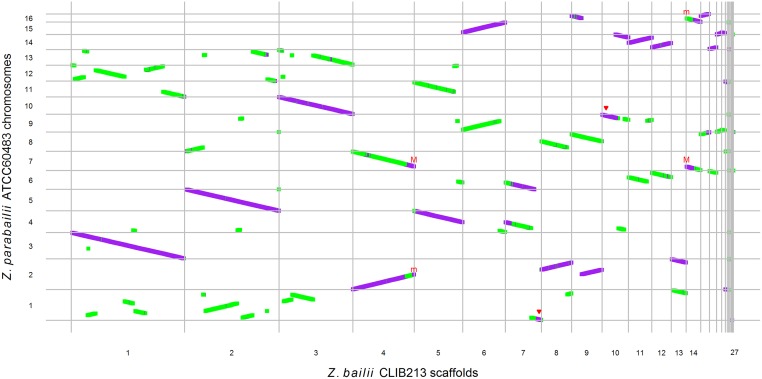
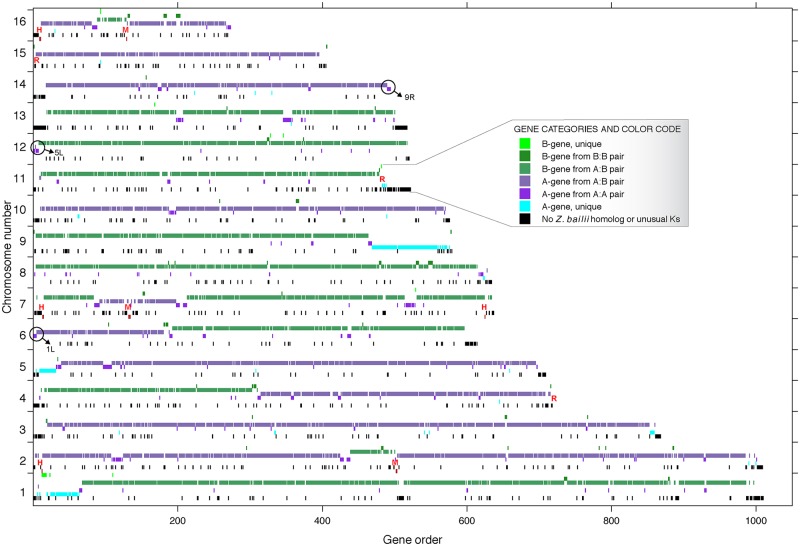
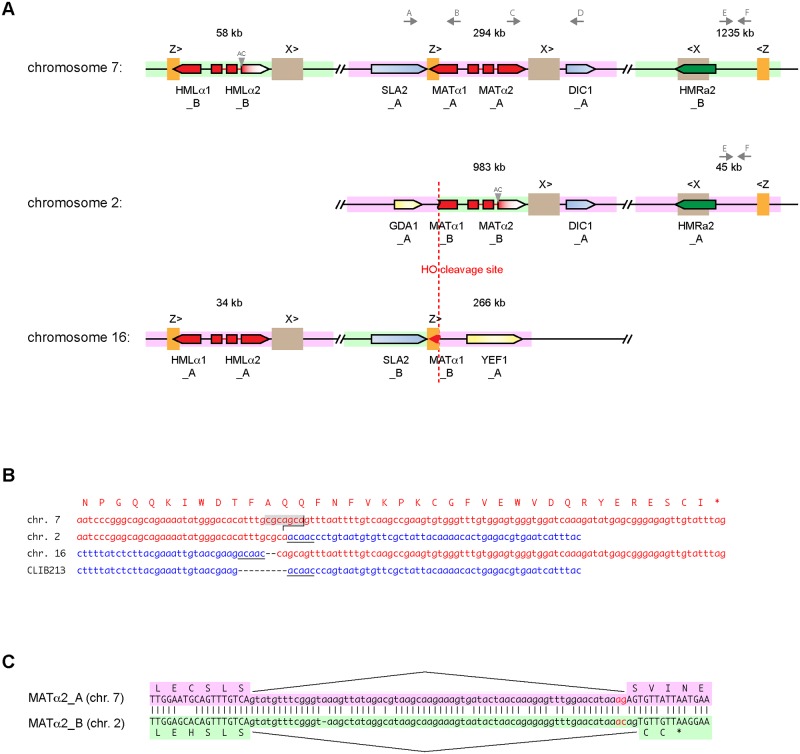
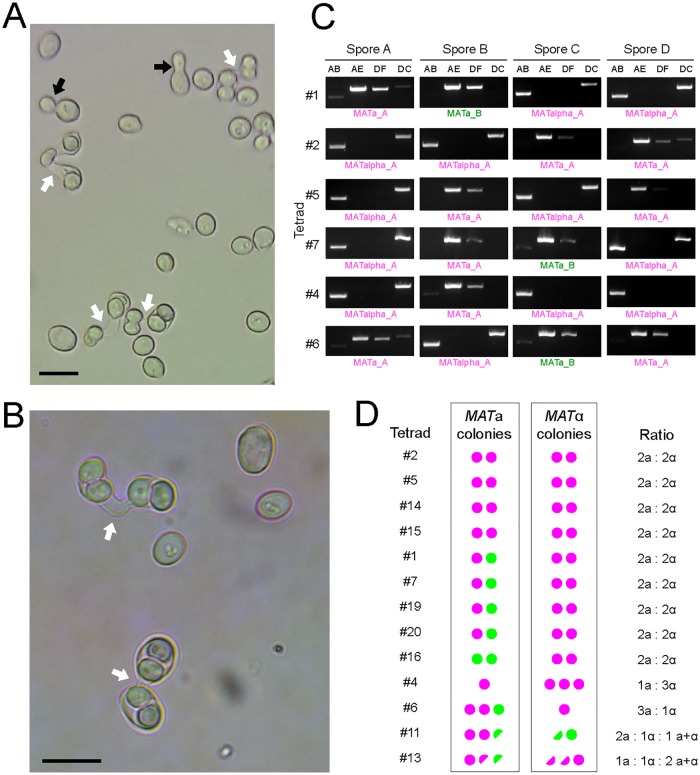
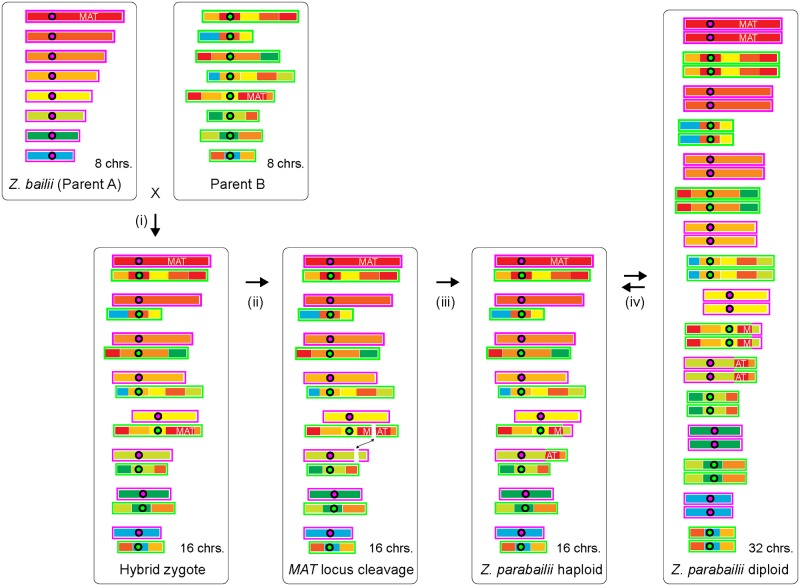
Many interspecies hybrids have been discovered in yeasts, but most of these hybrids are asexual and can replicate only mitotically. Whole-genome duplication has been proposed as a mechanism by which interspecies hybrids can regain fertility, restoring their ability to perform meiosis and sporulate. Here, we show that this process occurred naturally during the evolution of Zygosaccharomyces parabailii, an interspecies hybrid that was formed by mating between 2 parents that differed by 7% in genome sequence and by many interchromosomal rearrangements. Surprisingly, Z. parabailii has a full sexual cycle and is genetically haploid. It goes through mating-type switching and autodiploidization, followed by immediate sporulation. We identified the key evolutionary event that enabled Z. parabailii to regain fertility, which was breakage of 1 of the 2 homeologous copies of the mating-type (MAT) locus in the hybrid, resulting in a chromosomal rearrangement and irreparable damage to 1 MAT locus. This rearrangement was caused by HO endonuclease, which normally functions in mating-type switching. With 1 copy of MAT inactivated, the interspecies hybrid now behaves as a haploid. Our results provide the first demonstration that MAT locus damage is a naturally occurring evolutionary mechanism for whole-genome duplication and restoration of fertility to interspecies hybrids. The events that occurred in Z. parabailii strongly resemble those postulated to have caused ancient whole-genome duplication in an ancestor of Saccharomyces cerevisiae.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
Zygosaccharomyces pseudobailii, another yeast interspecies hybrid that regained fertility by damaging one of its MAT loci.FEMS Yeast Res. 2018 Nov 1;18(7):foy079. doi: 10.1093/femsyr/foy079. FEMS Yeast Res. 2018. PMID: 30052970 Free PMC article.
-
Mechanism for Restoration of Fertility in Hybrid Zygosaccharomyces rouxii Generated by Interspecies Hybridization.Appl Environ Microbiol. 2017 Oct 17;83(21):e01187-17. doi: 10.1128/AEM.01187-17. Print 2017 Nov 1. Appl Environ Microbiol. 2017. PMID: 28842546 Free PMC article.
-
Mating-type switching in CBS 732T derived subcultures unveils potential genetic and phenotypic novelties in haploid Zygosaccharomyces rouxii.FEMS Microbiol Lett. 2018 Feb 1;365(2). doi: 10.1093/femsle/fnx263. FEMS Microbiol Lett. 2018. PMID: 29228275
-
An Evolutionary Perspective on Yeast Mating-Type Switching.Genetics. 2017 May;206(1):9-32. doi: 10.1534/genetics.117.202036. Genetics. 2017. PMID: 28476860 Free PMC article. Review.
-
Hybridization and the origin of new yeast lineages.FEMS Yeast Res. 2020 Aug 1;20(5):foaa040. doi: 10.1093/femsyr/foaa040. FEMS Yeast Res. 2020. PMID: 32658267 Free PMC article. Review.
Cited by
-
Evolution and molecular bases of reproductive isolation.Curr Opin Genet Dev. 2022 Oct;76:101952. doi: 10.1016/j.gde.2022.101952. Epub 2022 Jul 16. Curr Opin Genet Dev. 2022. PMID: 35849861 Free PMC article. Review.
-
Extreme genome diversity and cryptic speciation in a harmful algal-bloom-forming eukaryote.Curr Biol. 2023 Jun 5;33(11):2246-2259.e8. doi: 10.1016/j.cub.2023.05.003. Epub 2023 May 23. Curr Biol. 2023. PMID: 37224809 Free PMC article.
-
The extremely halotolerant black yeast Hortaea werneckii - a model for intraspecific hybridization in clonal fungi.IMA Fungus. 2019 Jul 8;10:10. doi: 10.1186/s43008-019-0007-5. eCollection 2019. IMA Fungus. 2019. PMID: 32647617 Free PMC article.
-
Investigation of Genetic Relationships Between Hanseniaspora Species Found in Grape Musts Revealed Interspecific Hybrids With Dynamic Genome Structures.Front Microbiol. 2020 Jan 15;10:2960. doi: 10.3389/fmicb.2019.02960. eCollection 2019. Front Microbiol. 2020. PMID: 32010076 Free PMC article.
-
Transcriptional Response to Lactic Acid Stress in the Hybrid Yeast Zygosaccharomyces parabailii.Appl Environ Microbiol. 2018 Feb 14;84(5):e02294-17. doi: 10.1128/AEM.02294-17. Print 2018 Mar 1. Appl Environ Microbiol. 2018. PMID: 29269498 Free PMC article.
References
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
