

Morphological, anatomical and physiological leaf traits of Q. ilex, P. latifolia, P. lentiscus, and M. communis and their response to Mediterranean climate stress factors
- PMID: 28510875
- PMCID: PMC5432955
- DOI: 10.1186/1999-3110-54-35
Morphological, anatomical and physiological leaf traits of Q. ilex, P. latifolia, P. lentiscus, and M. communis and their response to Mediterranean climate stress factors
Abstract
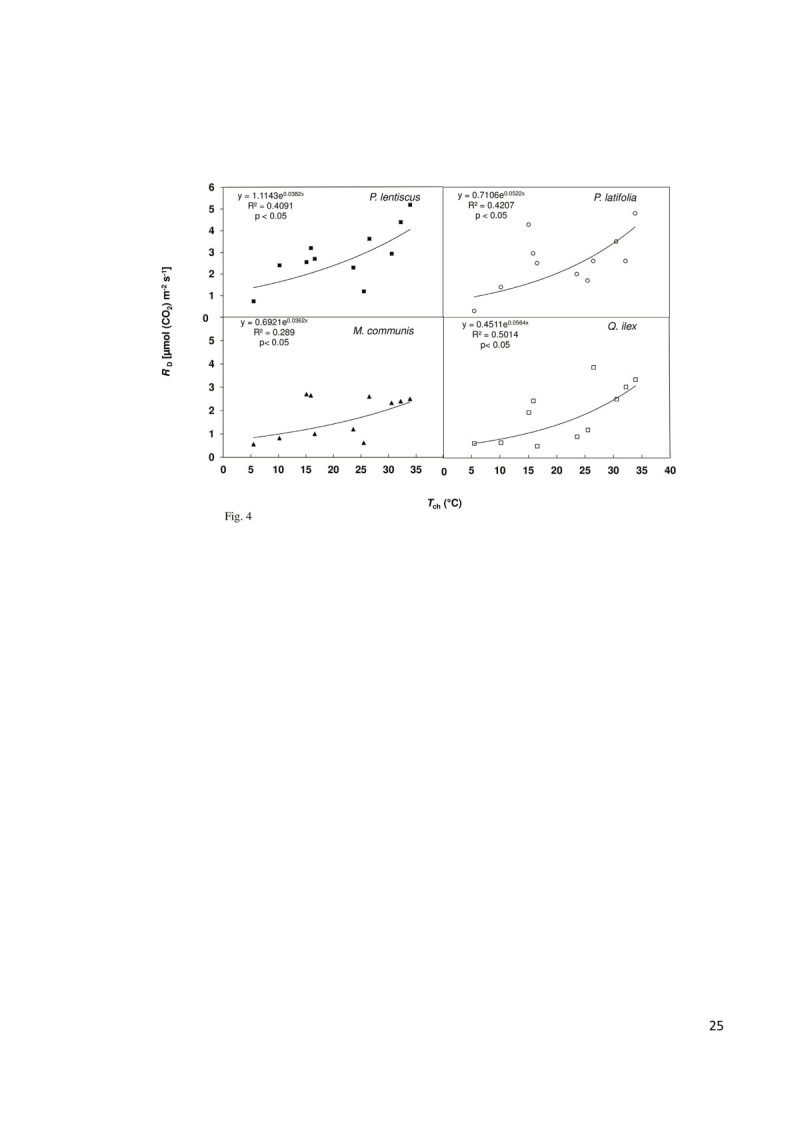
Background: Limitations to plant growth imposed by the Mediterranean climate are mainly due to carbon balance in response to stress factors. In particular, water stress associated to high air temperature and irradiance in summer causes a marked decrease in CO2 assimilation. Air temperature sensitivity of photosynthesis (PN) differs from that of leaf respiration (RD). PN often decreases sharply at temperature above its optimum while RD increases exponentially over short term rises in temperature. Nevertheless, the impact of water deficit on RD is still far from clear with reports in literature including decreases, maintenance or increases in its rates. The ratio RD/PN can be considered a simple approach to leaf carbon balance because it indicates the percentage of photosynthates that is respired.
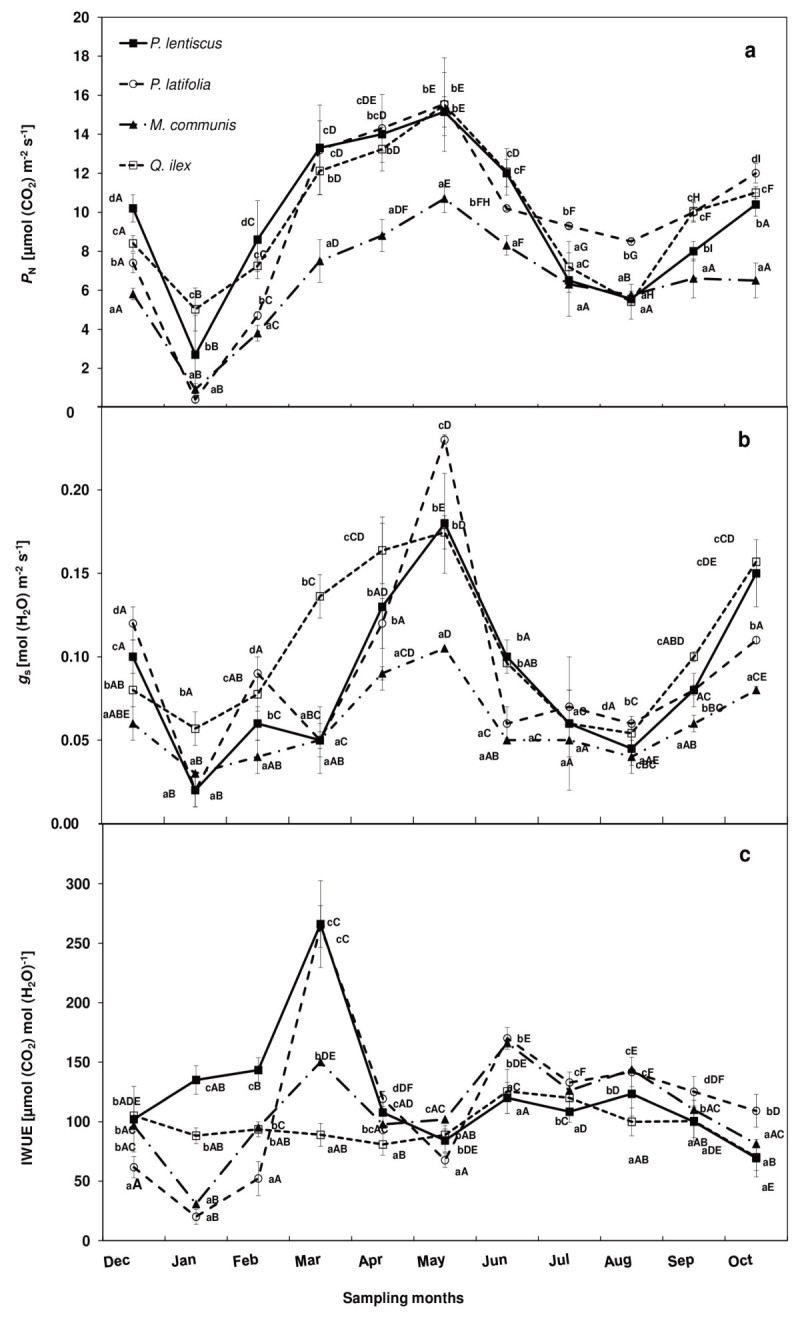
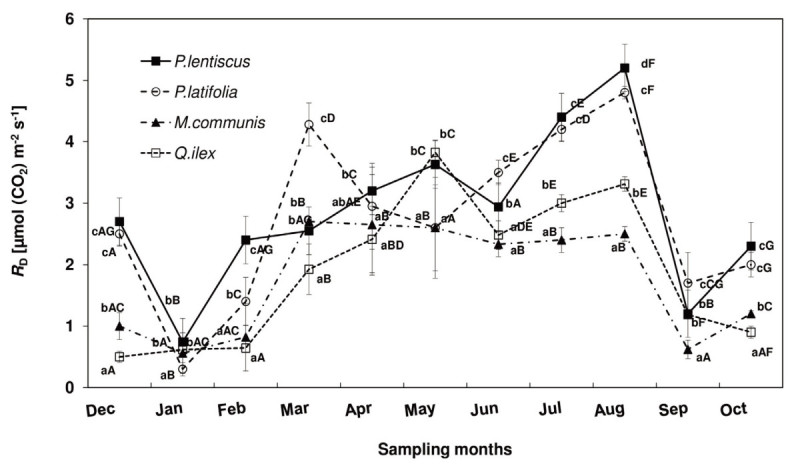
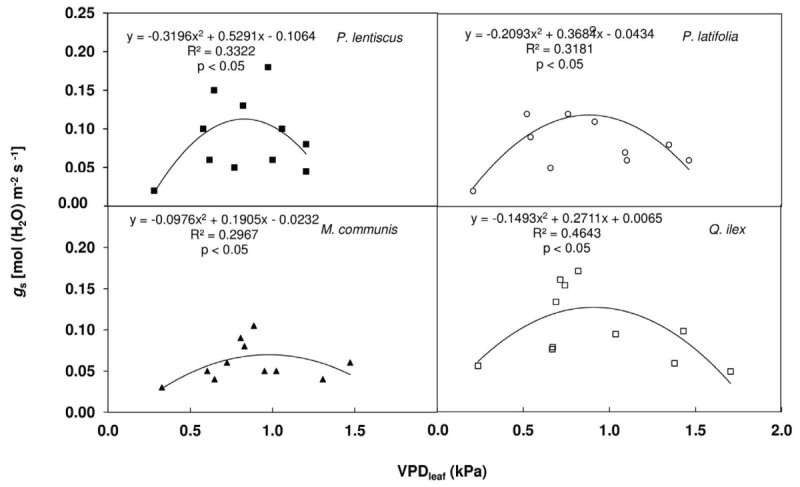
Results: The results underline different morphological, anatomical and physiological traits of the evergreen species co-occurring in the Mediterranean maquis which are indicative of their adaptive capability to Mediterranean stress factors. The ratio RD/PN varies from 0.15 ± 0.04 in autumn, 0.24 ± 0.05 in spring through 0.29 ± 0.15 in winter to 0.46 ± 0.11 in summer. The lower RD/PN in autumn and spring underlines the highest PN rates during the favorable periods when resources are not limited and leaves take in roughly three to five times more CO2 than they lose by respiration. On the contrary, the highest RD/PN ratio in summer underlines the lowest sensitivity of respiration to drought. Among the considered species, Quercus ilex and Pistacia lentiscus have the largest tolerance to low winter temperatures while Phillyrea latifolia and Myrtus communis to drought, and Phillyrea latifolia the highest recovery capability after the first rainfall following drought.
Conclusions: The Mediterranean evergreen specie shows a different tolerance to Mediterranean climate stress factors. The predicted global warming might differently affect carbon balance of the considered species, with a possible change in Mediterranean shrublands composition in the long-term. Understanding the carbon balance of plants in water limited environments is crucial in order to make informed land management decisions. Moreover, our results underline the importance of including seasonal variations of photosynthesis and respiration in carbon balance models.
Keywords: Air temperature; Drought; LMA; Leaf respiration; Myrtus communis; Phillyrea latifolia; Photosynthesis; Pistacia lentiscus; Quercus ilex.
Figures


Similar articles
-
Dampening effects of long-term experimental drought on growth and mortality rates of a Holm oak forest.Glob Chang Biol. 2013 Oct;19(10):3133-44. doi: 10.1111/gcb.12269. Epub 2013 Aug 8. Glob Chang Biol. 2013. PMID: 23712619
-
Responses to changes in Ca2+ supply in two Mediterranean evergreens, Phillyrea latifolia and Pistacia lentiscus, during salinity stress and subsequent relief.Ann Bot. 2008 Oct;102(4):609-22. doi: 10.1093/aob/mcn134. Epub 2008 Aug 13. Ann Bot. 2008. PMID: 18701601 Free PMC article.
-
Contrasting functional strategies following severe drought in two Mediterranean oaks with different leaf habit: Quercus faginea and Quercus ilex subsp. rotundifolia.Tree Physiol. 2021 Mar 6;41(3):371-387. doi: 10.1093/treephys/tpaa135. Tree Physiol. 2021. PMID: 33079165
-
Cavitation, stomatal conductance, and leaf dieback in seedlings of two co-occurring Mediterranean shrubs during an intense drought.J Exp Bot. 2003 Sep;54(390):2015-24. doi: 10.1093/jxb/erg221. Epub 2003 Jul 28. J Exp Bot. 2003. PMID: 12885857 Review.
-
Low leaf-level response to light and nutrients in Mediterranean evergreen oaks: a conservative resource-use strategy?New Phytol. 2000 Oct;148(1):79-91. doi: 10.1046/j.1469-8137.2000.00737.x. New Phytol. 2000. PMID: 33863045 Review.
Cited by
-
Genotype-specific responses to in vitro drought stress in myrtle (Myrtus communis L.): integrating machine learning techniques.PeerJ. 2024 Oct 7;12:e18081. doi: 10.7717/peerj.18081. eCollection 2024. PeerJ. 2024. PMID: 39391827 Free PMC article.
-
Seasonal, Sex- and Plant Size-Related Effects on Photoinhibition and Photoprotection in the Dioecious Mediterranean Dwarf Palm, Chamaerops humilis.Front Plant Sci. 2016 Jul 28;7:1116. doi: 10.3389/fpls.2016.01116. eCollection 2016. Front Plant Sci. 2016. PMID: 27516764 Free PMC article.
-
Seasonal and daily variations in primary and secondary metabolism of three maquis shrubs unveil different adaptive responses to Mediterranean climate.Conserv Physiol. 2019 Nov 5;7(1):coz070. doi: 10.1093/conphys/coz070. eCollection 2019. Conserv Physiol. 2019. PMID: 32467757 Free PMC article.
-
Study of functional and physiological response of co-occurring shrub species to the Mediterranean climate.Saudi J Biol Sci. 2019 Nov;26(7):1668-1675. doi: 10.1016/j.sjbs.2018.08.021. Epub 2018 Aug 23. Saudi J Biol Sci. 2019. PMID: 31762642 Free PMC article.
-
Profiling of Essential Oils from the Leaves of Pistacia lentiscus Collected in the Algerian Region of Tizi-Ouzou: Evidence of Chemical Variations Associated with Climatic Contrasts between Littoral and Mountain Samples.Molecules. 2022 Jun 28;27(13):4148. doi: 10.3390/molecules27134148. Molecules. 2022. PMID: 35807395 Free PMC article.
References
-
- Atkin OK, Evans JR, Ball MC, Siebke K. Relationship between the inhibition of leaf respiration by light and enhancement of leaf dark respiration following light treatment. Aust J Plant Physiol. 1998a;25:437–433. doi: 10.1071/PP97159. - DOI
-
- Atkin OK, Evans JR, Ball MC, Siebke K, Pons TL, Lambers H. Light inhibition of leaf respiration: the role of irradiance and temperature. In: Moller IM, Gardestrom P, Gliminius K, Glaser E, editors. Plant Mitochondria: from Gene to Function. Leiden: Backhuys Publishers; 1998b. pp. 567–574.
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
