

Tomato root microbiota and Phytophthora parasitica-associated disease
- PMID: 28511691
- PMCID: PMC5434524
- DOI: 10.1186/s40168-017-0273-7
Tomato root microbiota and Phytophthora parasitica-associated disease
Abstract
Background: Interactions between pathogenic oomycetes and microbiota residing on the surface of the host plant root are unknown, despite being critical to inoculum constitution. The nature of these interactions was explored for the polyphagous and telluric species Phytophthora parasitica.
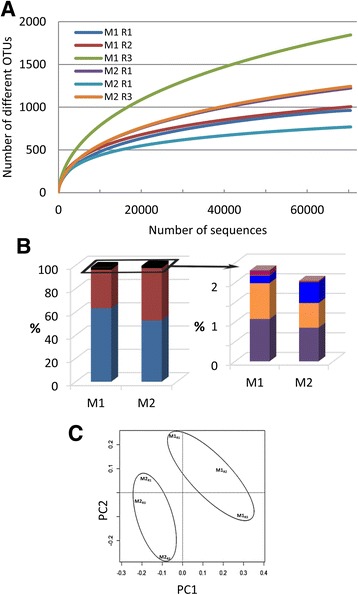
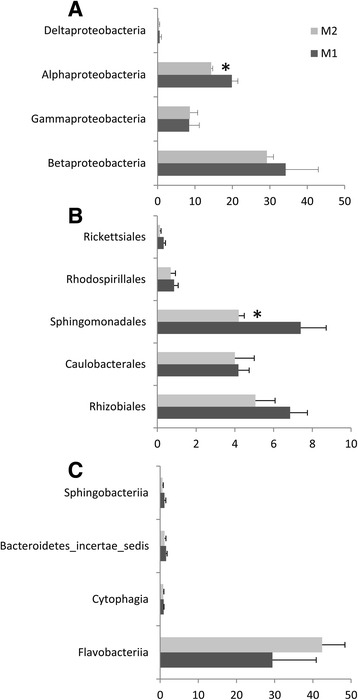
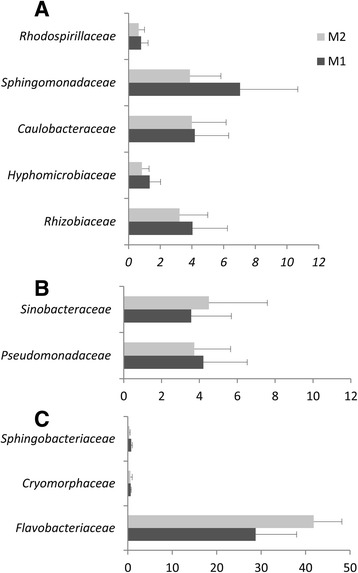
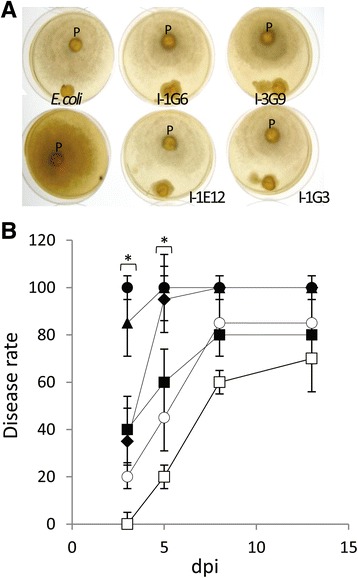
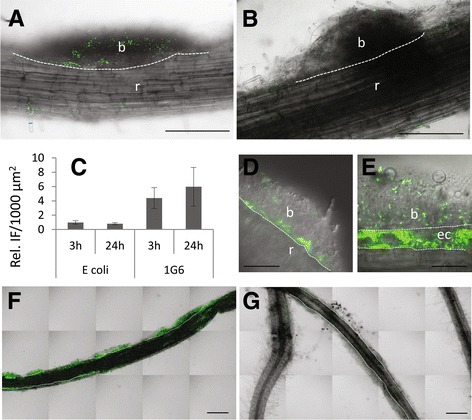
Results: Composition of the rhizospheric microbiota of Solanum lycopersicum was characterized using deep re-sequencing of 16S rRNA gene to analyze tomato roots either free of or partly covered with P. parasitica biofilm. Colonization of the host root surface by the oomycete was associated with a shift in microbial community involving a Bacteroidetes/Proteobacteria transition and Flavobacteriaceae as the most abundant family. Identification of members of the P. parasitica-associated microbiota interfering with biology and oomycete infection was carried out by screening for bacteria able to (i) grow on a P. parasitica extract-based medium (ii), exhibit in vitro probiotic or antibiotic activity towards the oomycete (iii), have an impact on the oomycete infection cycle in a tripartite interaction S. lycopersicum-P. parasitica-bacteria. One Pseudomonas phylotype was found to exacerbate disease symptoms in tomato plants. The lack of significant gene expression response of P. parasitica effectors to Pseudomonas suggested that the increase in plant susceptibility was not associated with an increase in virulence. Our results reveal that Pseudomonas spp. establishes commensal interactions with the oomycete. Bacteria preferentially colonize the surface of the biofilm rather than the roots, so that they can infect plant cells without any apparent infection of P. parasitica.
Conclusions: The presence of the pathogenic oomycete P. parasitica in the tomato rhizosphere leads to a shift in the rhizospheric microbiota composition. It contributes to the habitat extension of Pseudomonas species mediated through a physical association between the oomycete and the bacteria.
Keywords: Biofilm; Flavobacteriaceae; Host plant; Metagenomics 16S; Oomycete; Pseudomonadaceae.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
