

Glutathionylation of Yersinia pestis LcrV and Its Effects on Plague Pathogenesis
- PMID: 28512097
- PMCID: PMC5433101
- DOI: 10.1128/mBio.00646-17
Glutathionylation of Yersinia pestis LcrV and Its Effects on Plague Pathogenesis
Abstract
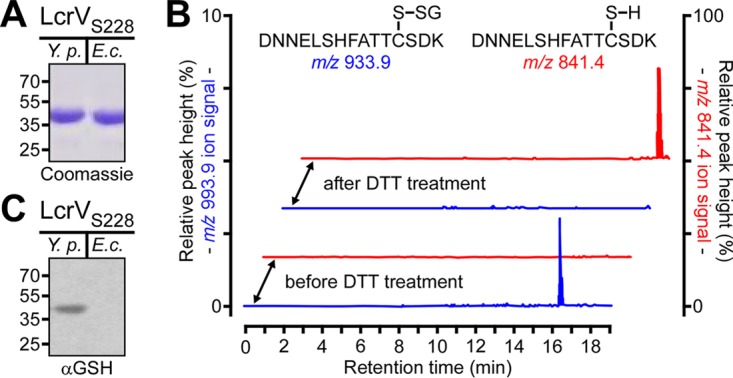
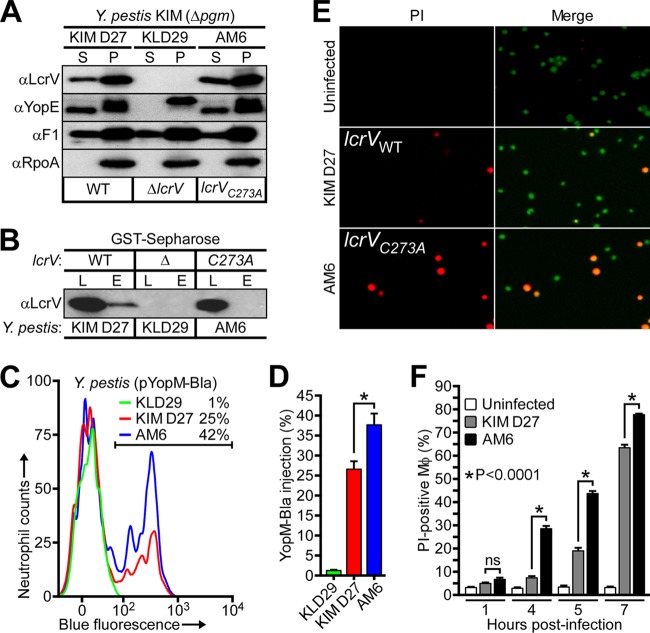
Glutathionylation, the formation of reversible mixed disulfides between glutathione and protein cysteine residues, is a posttranslational modification previously observed for intracellular proteins of bacteria. Here we show that Yersinia pestis LcrV, a secreted protein capping the type III secretion machine, is glutathionylated at Cys273 and that this modification promotes association with host ribosomal protein S3 (RPS3), moderates Y. pestis type III effector transport and killing of macrophages, and enhances bubonic plague pathogenesis in mice and rats. Secreted LcrV was purified and analyzed by mass spectrometry to reveal glutathionylation, a modification that is abolished by the codon substitution Cys273Ala in lcrV Moreover, the lcrVC273A mutation enhanced the survival of animals in models of bubonic plague. Investigating the molecular mechanism responsible for these virulence attributes, we identified macrophage RPS3 as a ligand of LcrV, an association that is perturbed by the Cys273Ala substitution. Furthermore, macrophages infected by the lcrVC273A variant displayed accelerated apoptotic death and diminished proinflammatory cytokine release. Deletion of gshB, which encodes glutathione synthetase of Y. pestis, resulted in undetectable levels of intracellular glutathione, and we used a Y. pestis ΔgshB mutant to characterize the biochemical pathway of LcrV glutathionylation, establishing that LcrV is modified after its transport to the type III needle via disulfide bond formation with extracellular oxidized glutathione.IMPORTANCEYersinia pestis, the causative agent of plague, has killed large segments of the human population; however, the molecular bases for the extraordinary virulence attributes of this pathogen are not well understood. We show here that LcrV, the cap protein of bacterial type III secretion needles, is modified by host glutathione and that this modification contributes to the high virulence of Y. pestis in mouse and rat models for bubonic plague. These data suggest that Y. pestis exploits glutathione in host tissues to activate a virulence strategy, thereby accelerating plague pathogenesis.
Keywords: Yersinia pestis; glutathionylation; innate immunity; macrophage; plague; ribosomal protein S3 (RPS3).
Copyright © 2017 Mitchell et al.
Figures
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
