

Divergent Requirements for EZH1 in Heart Development Versus Regeneration
- PMID: 28512107
- PMCID: PMC5527745
- DOI: 10.1161/CIRCRESAHA.117.311212
Divergent Requirements for EZH1 in Heart Development Versus Regeneration
Abstract
Rationale: Polycomb repressive complex 2 is a major epigenetic repressor that deposits methylation on histone H3 on lysine 27 (H3K27me) and controls differentiation and function of many cells, including cardiac myocytes. EZH1 and EZH2 are 2 alternative catalytic subunits with partial functional redundancy. The relative roles of EZH1 and EZH2 in heart development and regeneration are unknown.
Objective: We compared the roles of EZH1 versus EZH2 in heart development and neonatal heart regeneration.
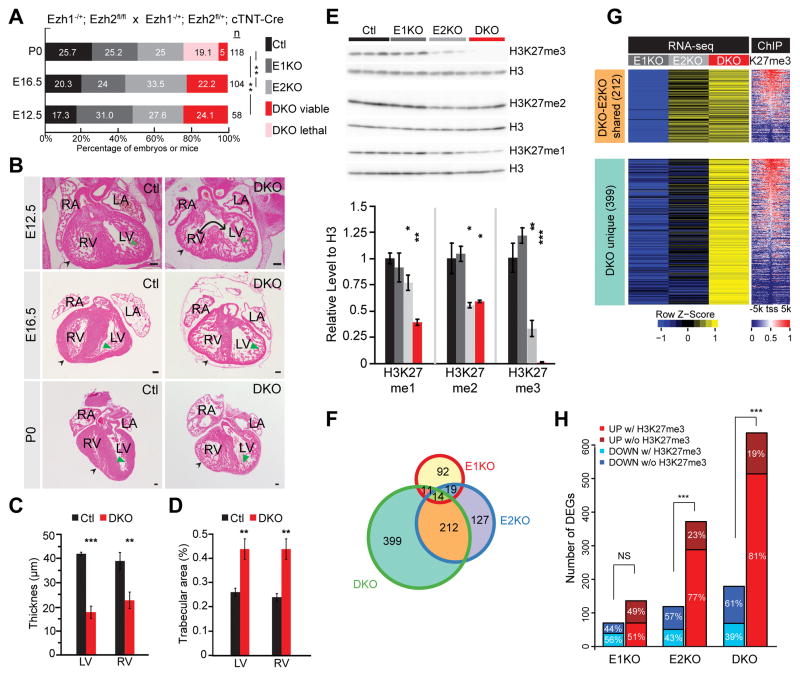
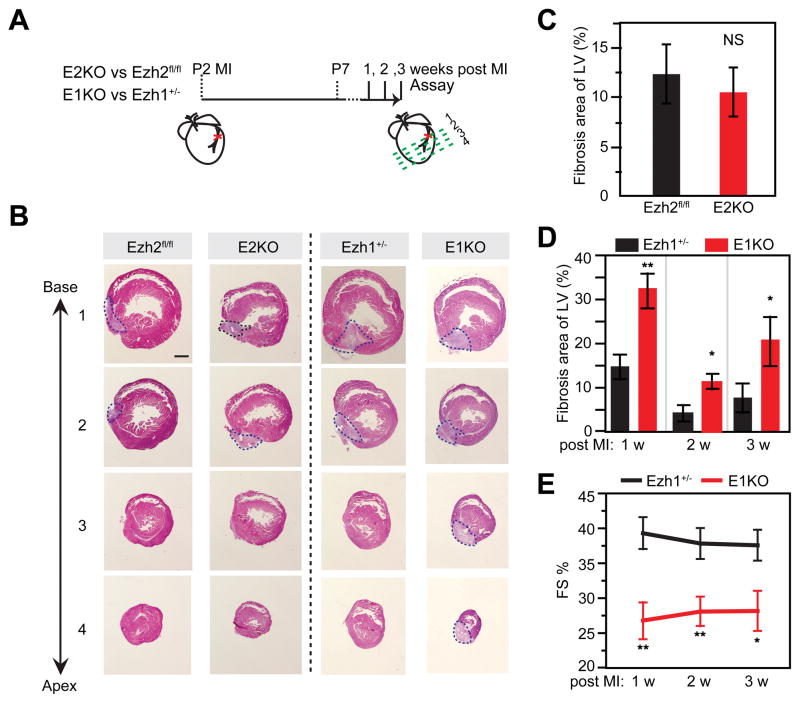
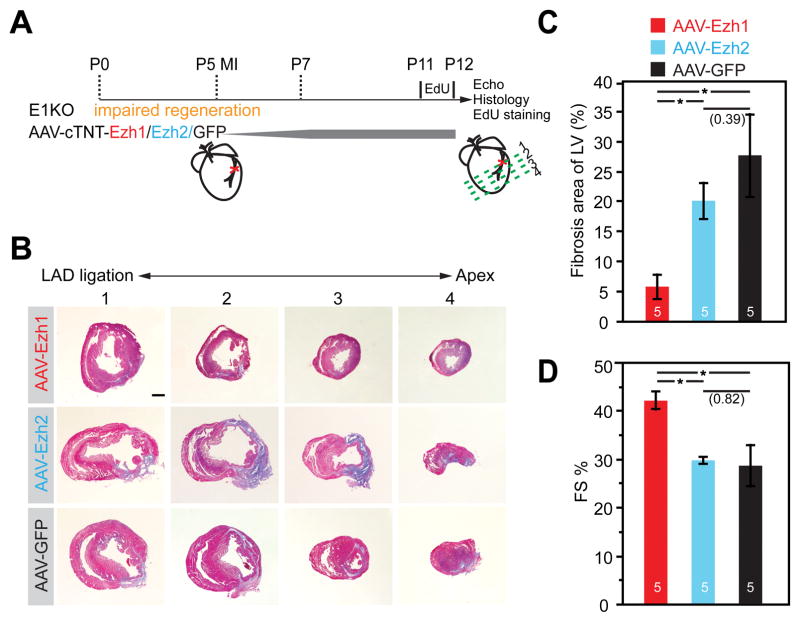
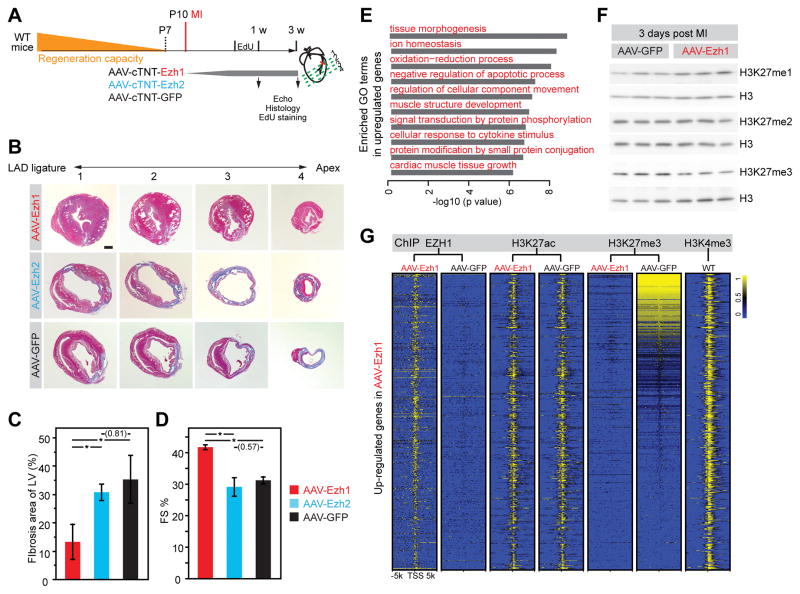
Methods and results: Heart development was normal in Ezh1-/- (Ezh1 knockout) and Ezh2f/f::cTNT-Cre (Ezh2 knockout) embryos. Ablation of both genes in Ezh1-/-::Ezh2f/f::cTNT-Cre embryos caused lethal heart malformations, including hypertrabeculation, compact myocardial hypoplasia, and ventricular septal defect. Epigenome and transcriptome profiling showed that derepressed genes were upregulated in a manner consistent with total EZH dose. In neonatal heart regeneration, Ezh1 was required, but Ezh2 was dispensable. This finding was further supported by rescue experiments: cardiac myocyte-restricted re-expression of EZH1 but not EZH2 restored neonatal heart regeneration in Ezh1 knockout. In myocardial infarction performed outside of the neonatal regenerative window, EZH1 but not EZH2 likewise improved heart function and stimulated cardiac myocyte proliferation. Mechanistically, EZH1 occupied and activated genes related to cardiac growth.
Conclusions: Our work unravels divergent mechanisms of EZH1 in heart development and regeneration, which will empower efforts to overcome epigenetic barriers to heart regeneration.
Keywords: lysine; methylation; myocardium; polycomb repressive complex 2; regeneration.
© 2017 American Heart Association, Inc.
Conflict of interest statement
The authors have no conflicts of interest to disclose.
Figures
Comment in
-
EZ Switch From EZH2 to EZH1: Histone Methylation Opens a Window of Cardiac Regeneration.Circ Res. 2017 Jul 7;121(2):91-94. doi: 10.1161/CIRCRESAHA.117.311351. Circ Res. 2017. PMID: 28684617 No abstract available.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
