

Cap-n-Collar Promotes Tissue Regeneration by Regulating ROS and JNK Signaling in the Drosophila melanogaster Wing Imaginal Disc
- PMID: 28512185
- PMCID: PMC5500147
- DOI: 10.1534/genetics.116.196832
Cap-n-Collar Promotes Tissue Regeneration by Regulating ROS and JNK Signaling in the Drosophila melanogaster Wing Imaginal Disc
Abstract
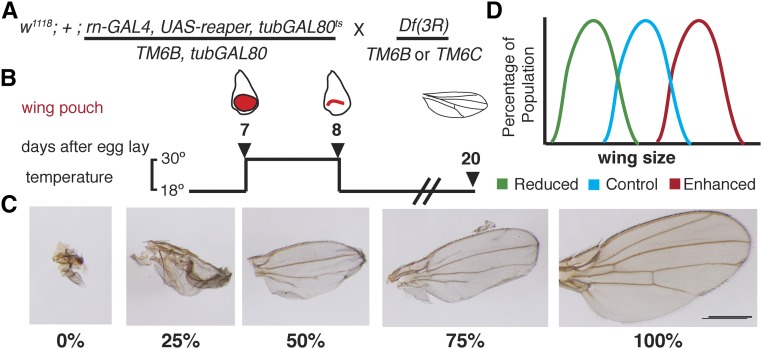
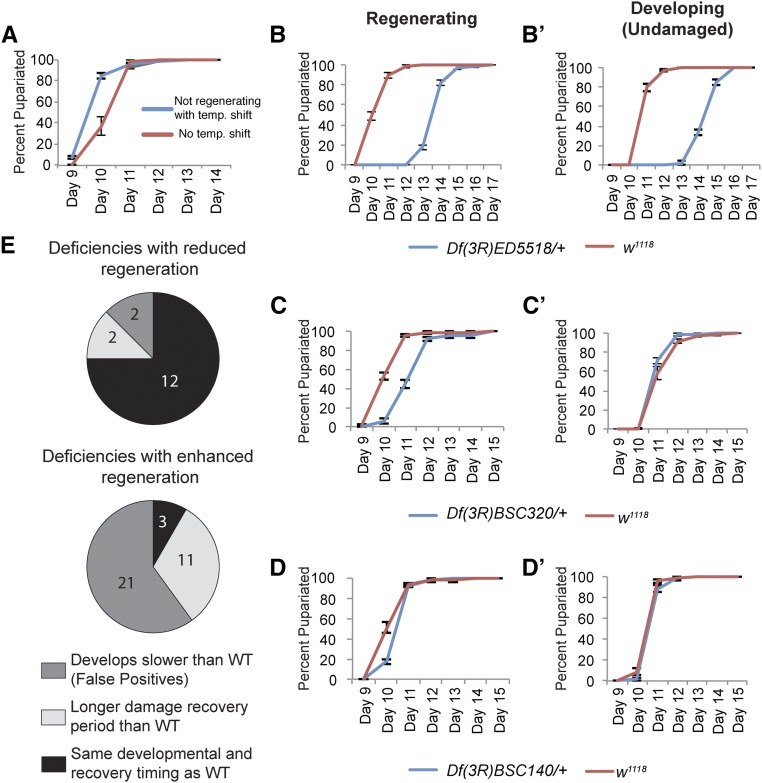
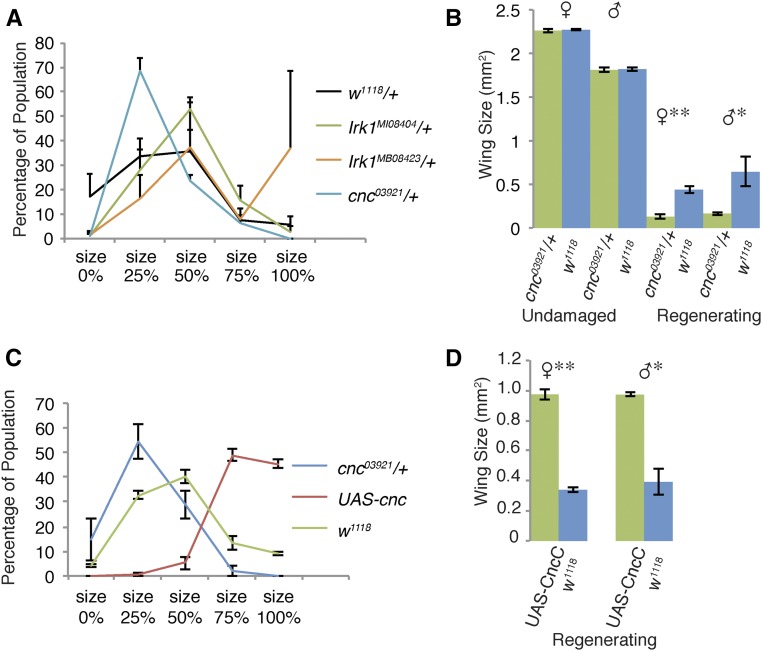
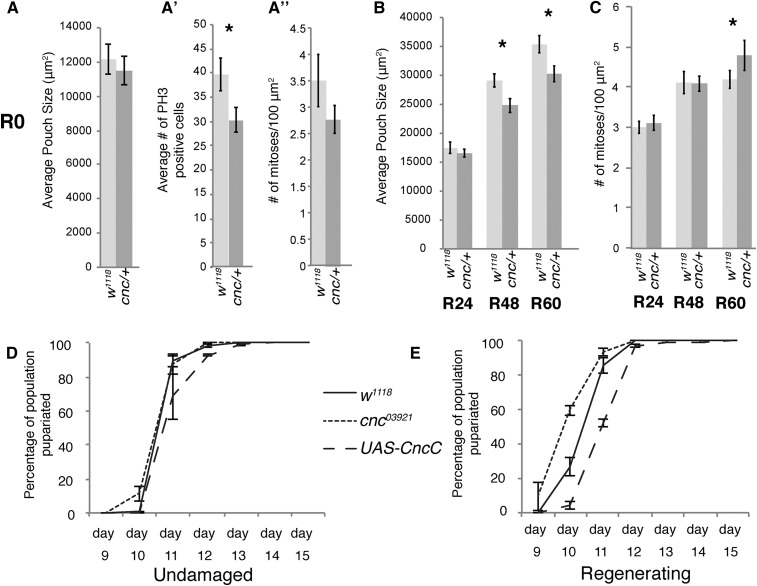
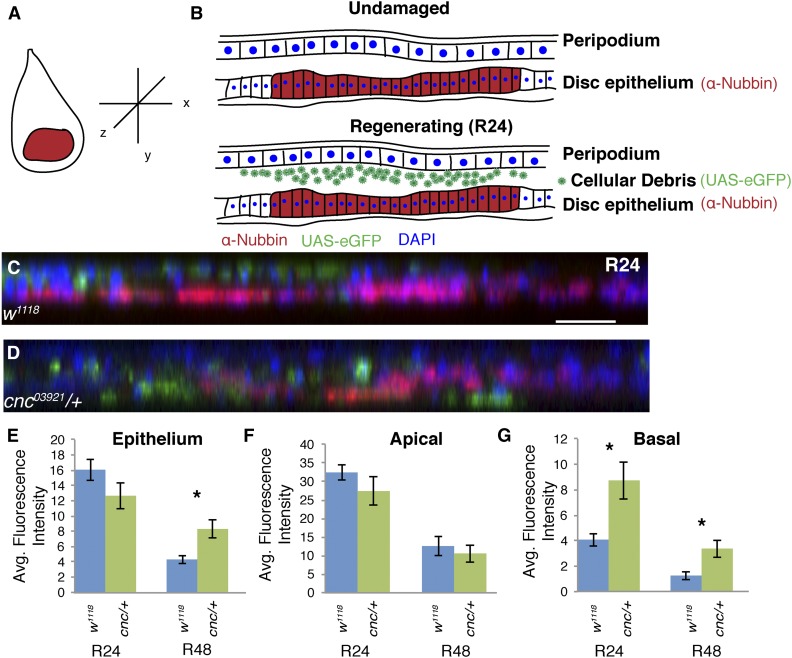
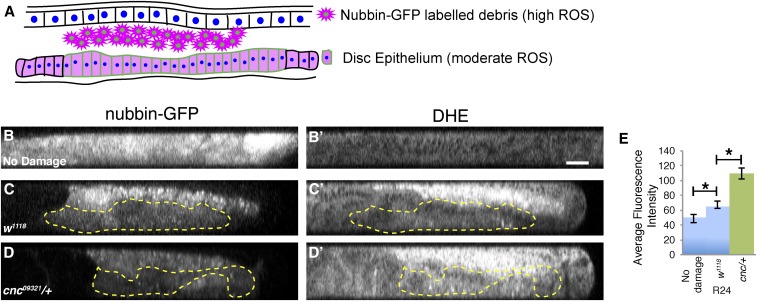
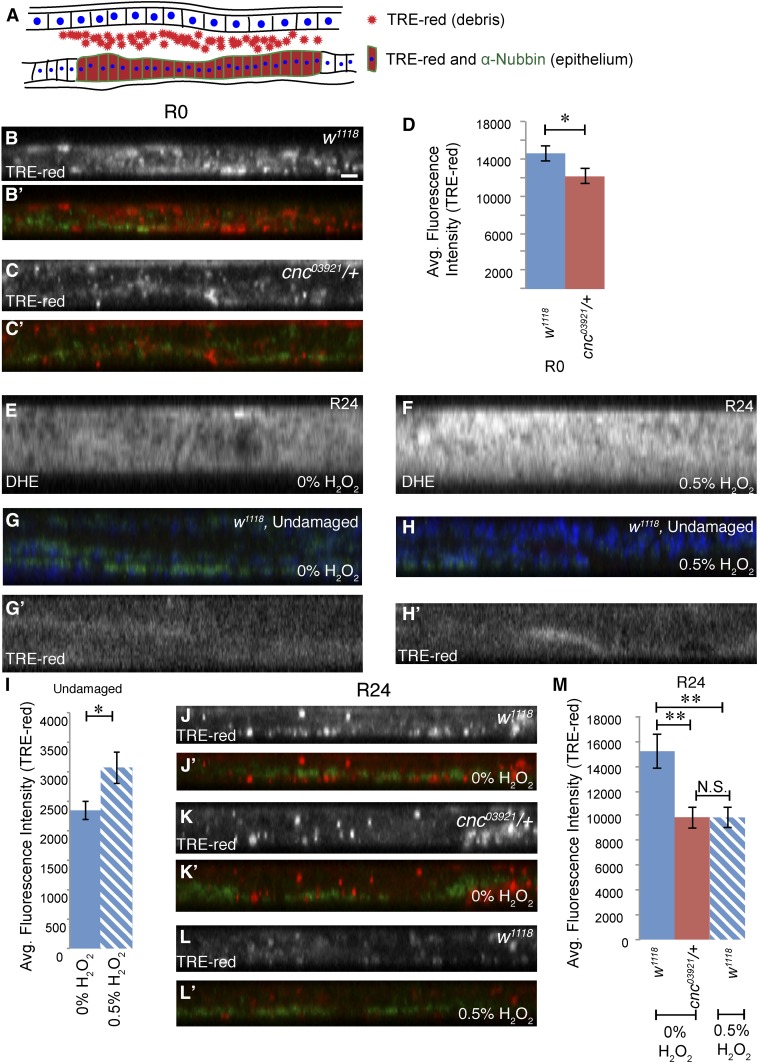
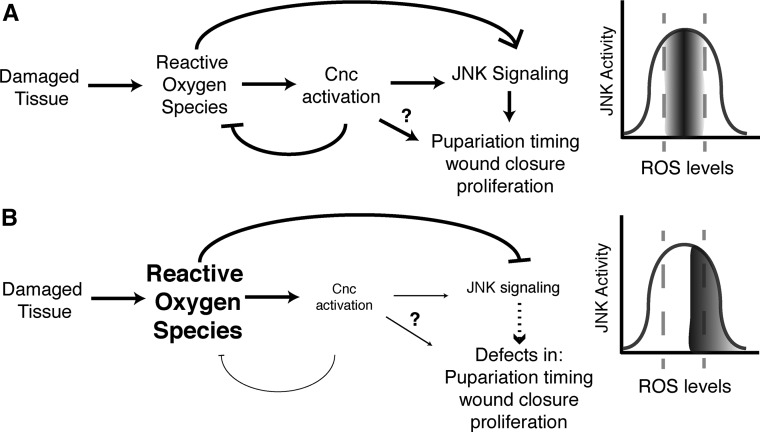
Regeneration is a complex process that requires an organism to recognize and repair tissue damage, as well as grow and pattern new tissue. Here, we describe a genetic screen to identify novel regulators of regeneration. We ablated the Drosophila melanogaster larval wing primordium by inducing apoptosis in a spatially and temporally controlled manner and allowed the tissue to regenerate and repattern. To identify genes that regulate regeneration, we carried out a dominant-modifier screen by assessing the amount and quality of regeneration in adult wings heterozygous for isogenic deficiencies. We have identified 31 regions on the right arm of the third chromosome that modify the regenerative response. Interestingly, we observed several distinct phenotypes: mutants that regenerated poorly, mutants that regenerated faster or better than wild-type, and mutants that regenerated imperfectly and had patterning defects. We mapped one deficiency region to cap-n-collar (cnc), the Drosophila Nrf2 ortholog, which is required for regeneration. Cnc regulates reactive oxygen species levels in the regenerating epithelium, and affects c-Jun N-terminal protein kinase (JNK) signaling, growth, debris localization, and pupariation timing. Here, we present the results of our screen and propose a model wherein Cnc regulates regeneration by maintaining an optimal level of reactive oxygen species to promote JNK signaling.
Keywords: Cap-n-collar; Drosophila; Nrf2; imaginal disc; reactive oxygen species; regeneration.
Copyright © 2017 by the Genetics Society of America.
Figures
References
-
- Bando T., Mito T., Maeda Y., Nakamura T., Ito F., et al. , 2009. Regulation of leg size and shape by the Dachsous/Fat signalling pathway during regeneration. Development 136: 2235–2245. - PubMed
-
- Bergantiños C., Corominas M., Serras F., 2010. Cell death-induced regeneration in wing imaginal discs requires JNK signalling. Development 137: 1169–1179. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
