

Non-homologous DNA end joining and alternative pathways to double-strand break repair
- PMID: 28512351
- PMCID: PMC7062608
- DOI: 10.1038/nrm.2017.48
Non-homologous DNA end joining and alternative pathways to double-strand break repair
Abstract
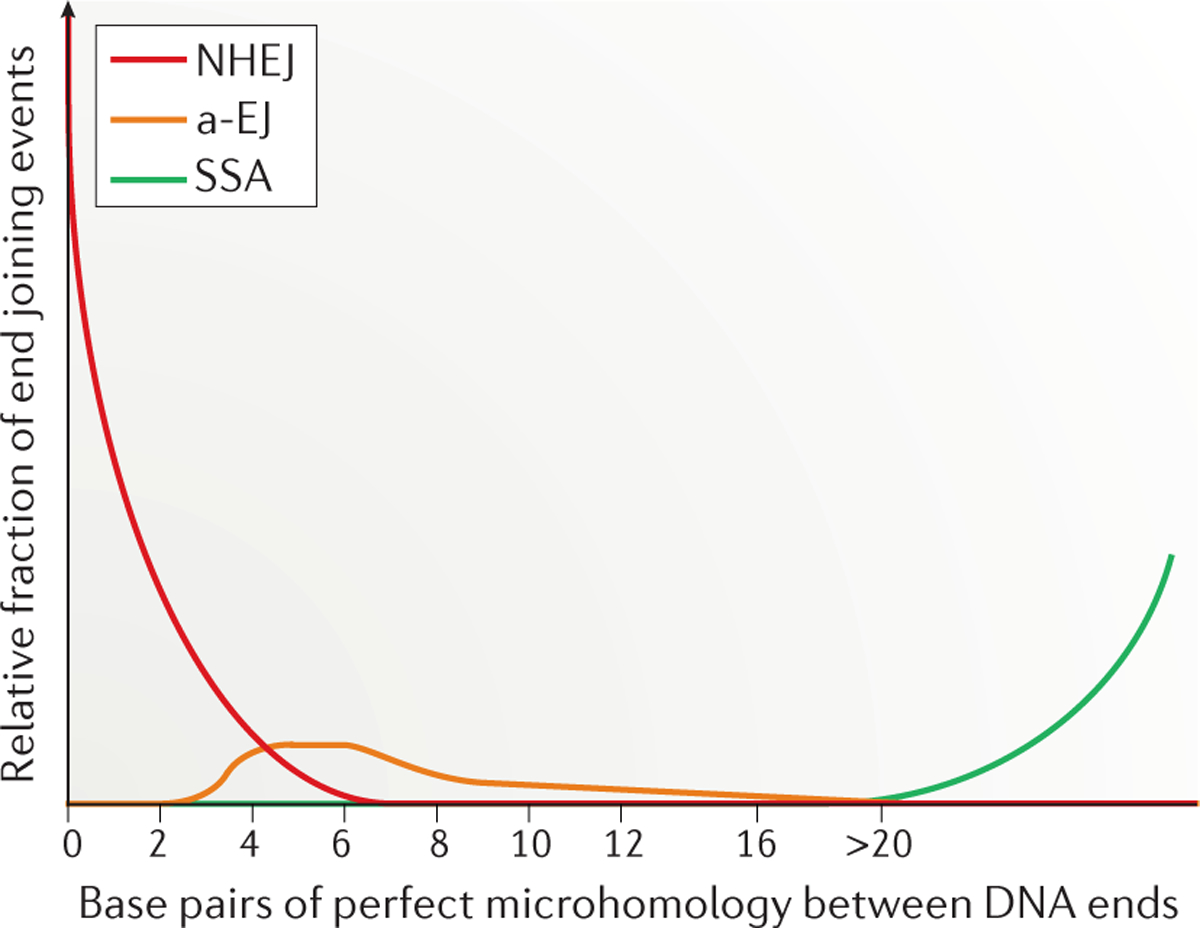
DNA double-strand breaks (DSBs) are the most dangerous type of DNA damage because they can result in the loss of large chromosomal regions. In all mammalian cells, DSBs that occur throughout the cell cycle are repaired predominantly by the non-homologous DNA end joining (NHEJ) pathway. Defects in NHEJ result in sensitivity to ionizing radiation and the ablation of lymphocytes. The NHEJ pathway utilizes proteins that recognize, resect, polymerize and ligate the DNA ends in a flexible manner. This flexibility permits NHEJ to function on a wide range of DNA-end configurations, with the resulting repaired DNA junctions often containing mutations. In this Review, we discuss the most recent findings regarding the relative involvement of the different NHEJ proteins in the repair of various DNA-end configurations. We also discuss the shunting of DNA-end repair to the auxiliary pathways of alternative end joining (a-EJ) or single-strand annealing (SSA) and the relevance of these different pathways to human disease.
Conflict of interest statement
Competing interests statement
The authors declare no competing interests.
Figures
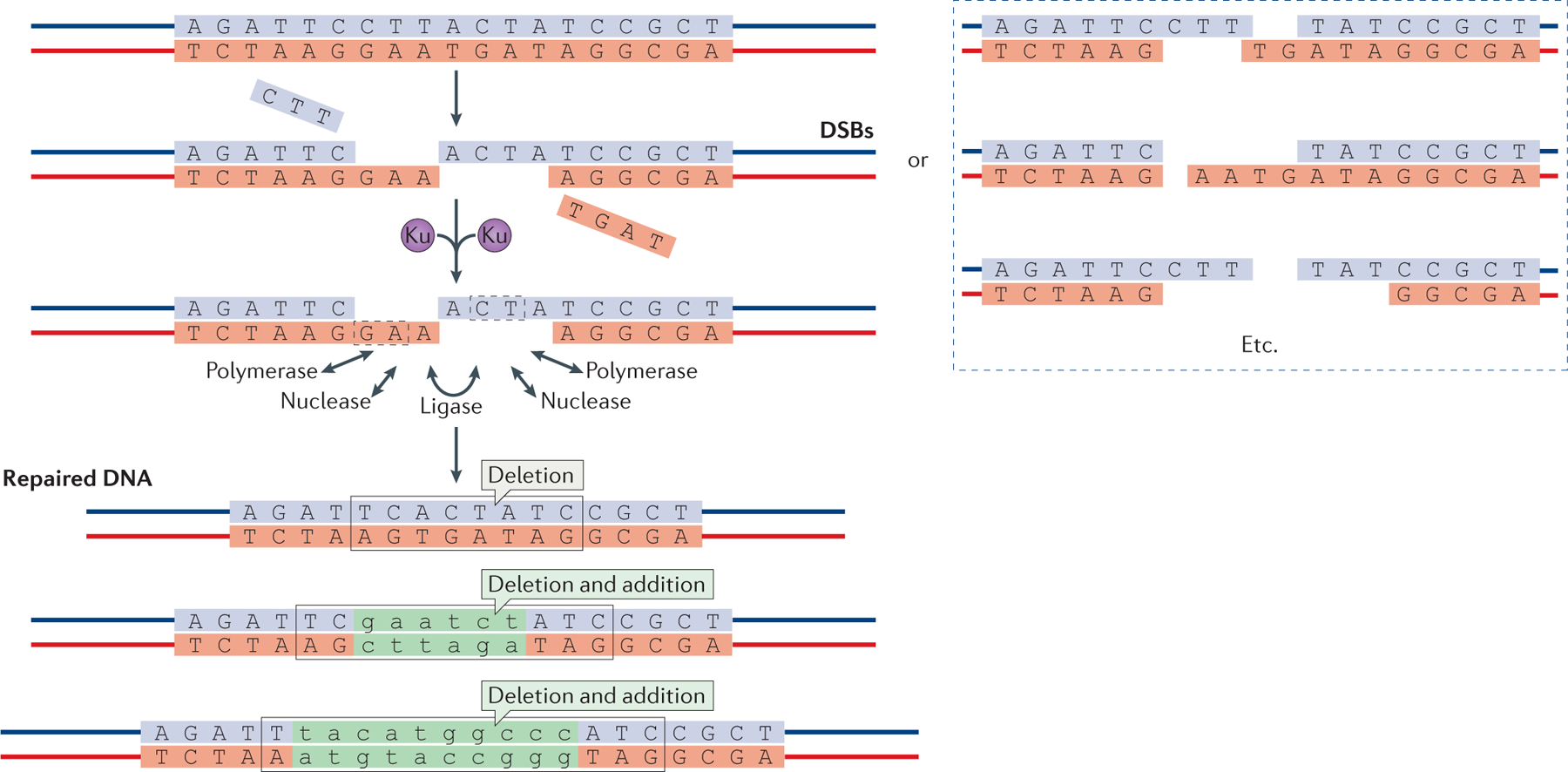
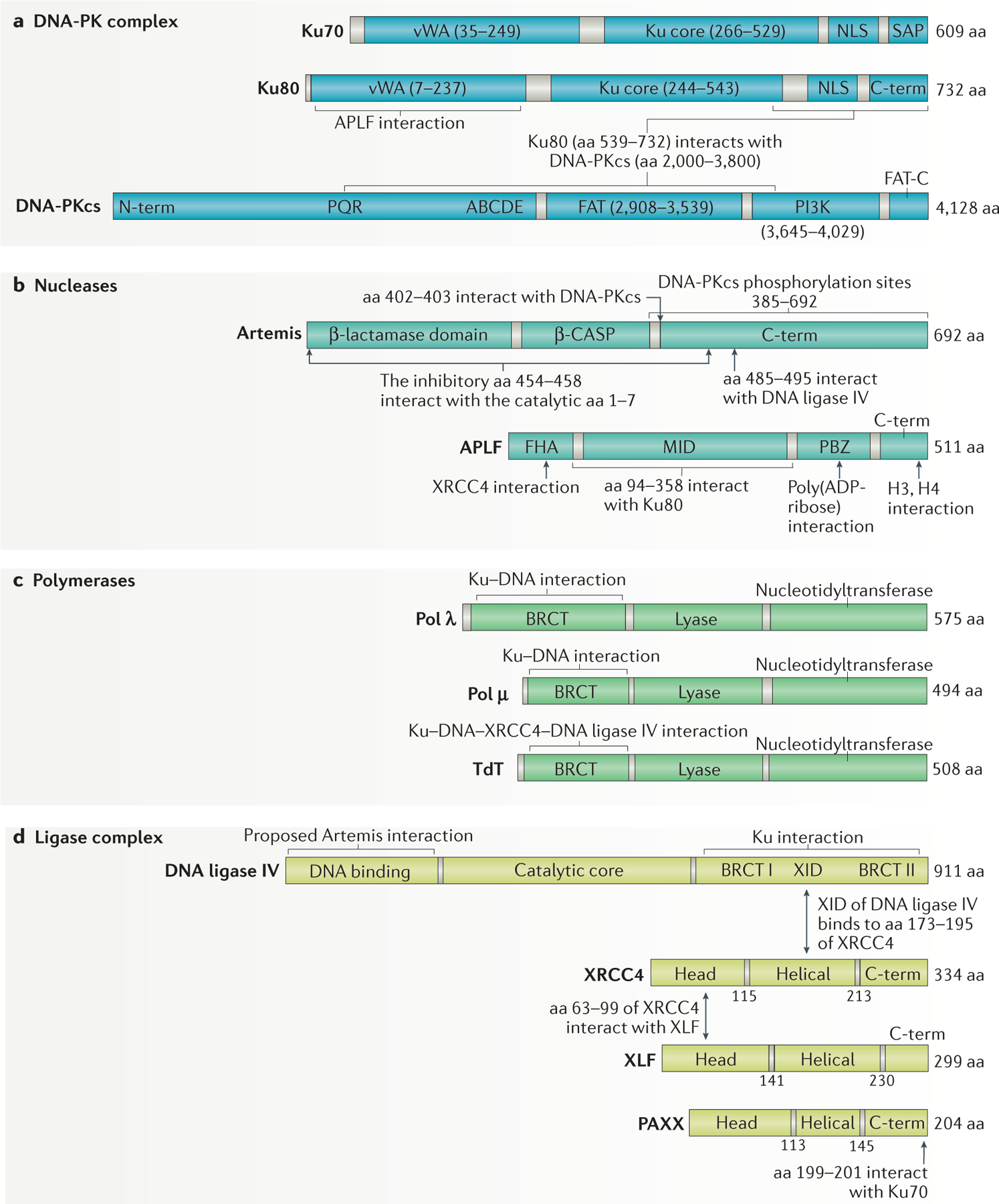
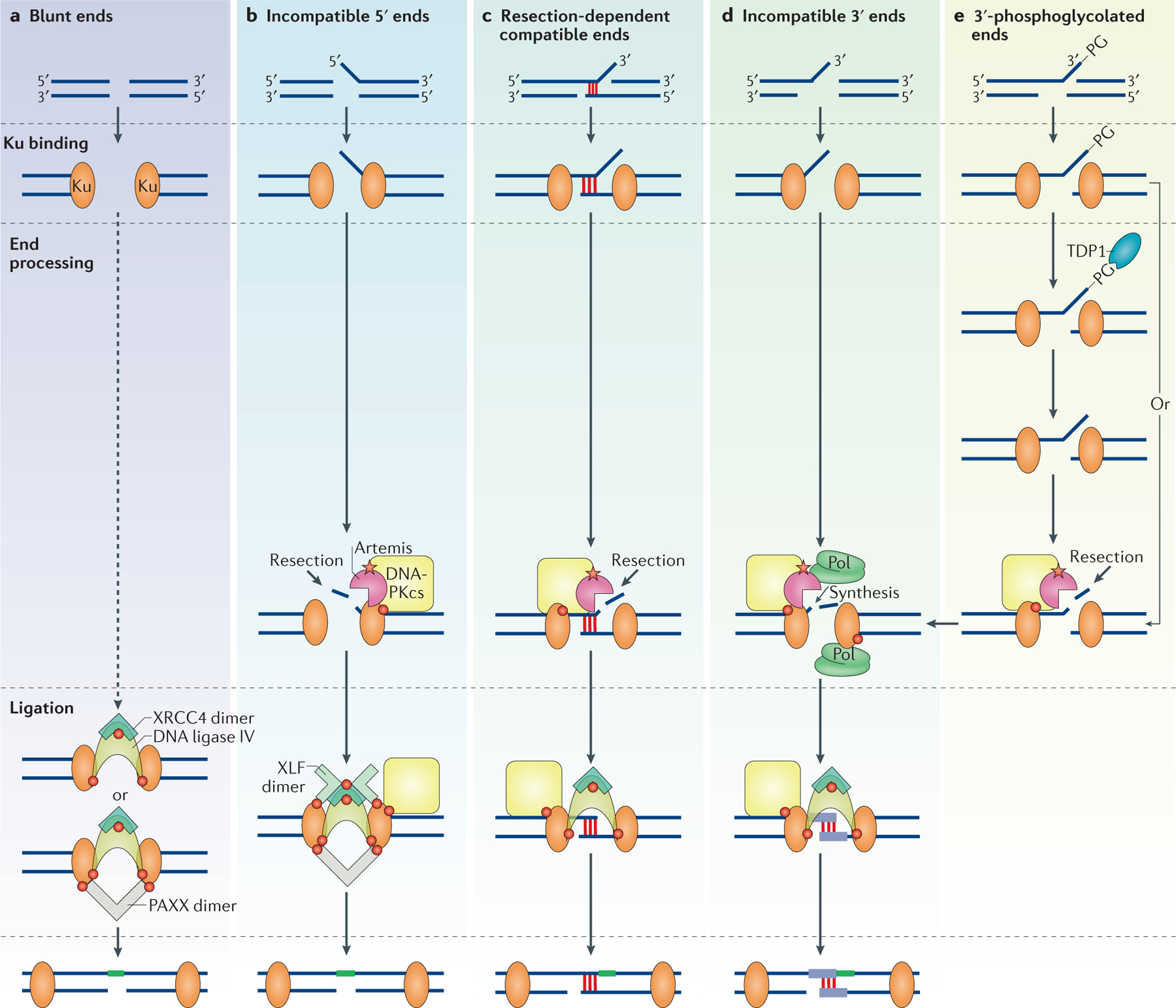
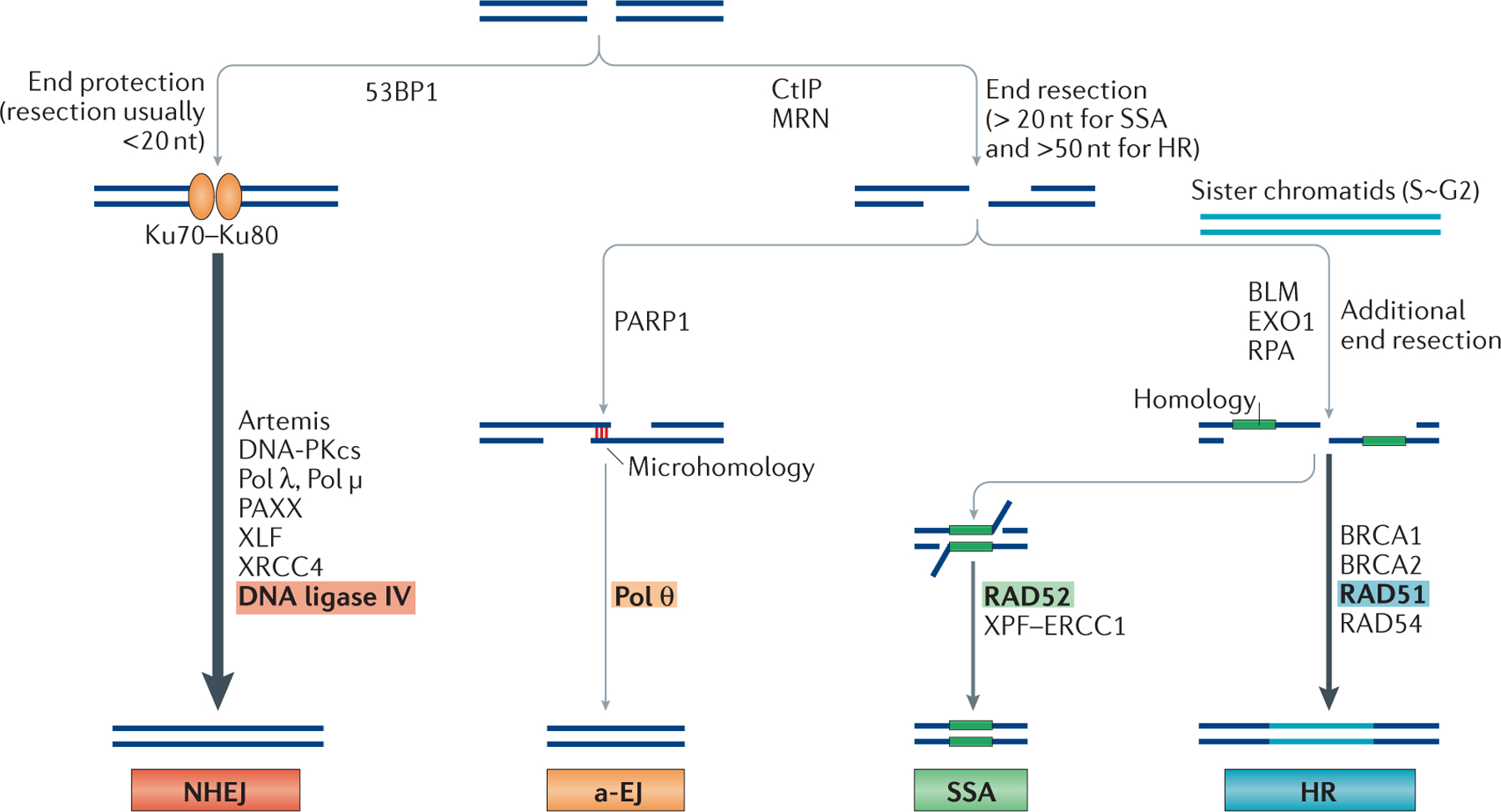
References
-
- Lieber MR & Karanjawala ZE Ageing, repetitive genomes and DNA damage. Nat. Rev. Mol. Cell Biol 5, 69–75 (2004). - PubMed
-
- Martin GM, Smith AC, Ketterer DJ, Ogburn CE & Disteche CM Increased chromosomal aberrations in first metaphases of cells isolated from the kidneys of aged mice. Isr. J. Med. Sci 21, 296–301 (1985). - PubMed
-
- Meek K, Dang V & Lees-Miller SP DNA-PK: the means to justify the ends? Adv. Immunol 99, 33–58 (2008). - PubMed
-
- Chang HHY et al. Different DNA end configurations dictate which NHEJ components are most important for joining efficiency. J. Biol. Chem 291, 24377–24389 (2016). - PMC - PubMed
-
This paper describes a biochemical reconstitution of NHEJ using a direct gel assay (without PCR) and all of the major NHEJ components.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
