

APC/C-Cdc20 mediates deprotection of centromeric cohesin at meiosis II in yeast
- PMID: 28514186
- PMCID: PMC5499901
- DOI: 10.1080/15384101.2017.1320628
APC/C-Cdc20 mediates deprotection of centromeric cohesin at meiosis II in yeast
Abstract
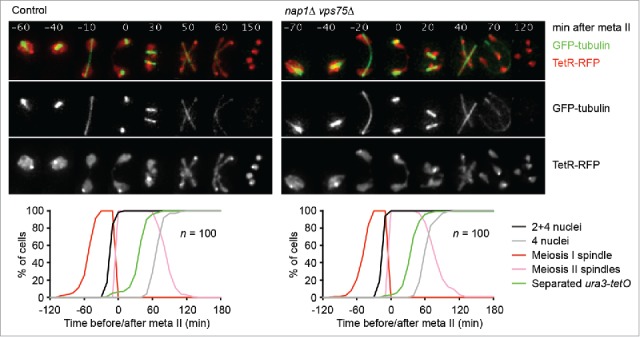
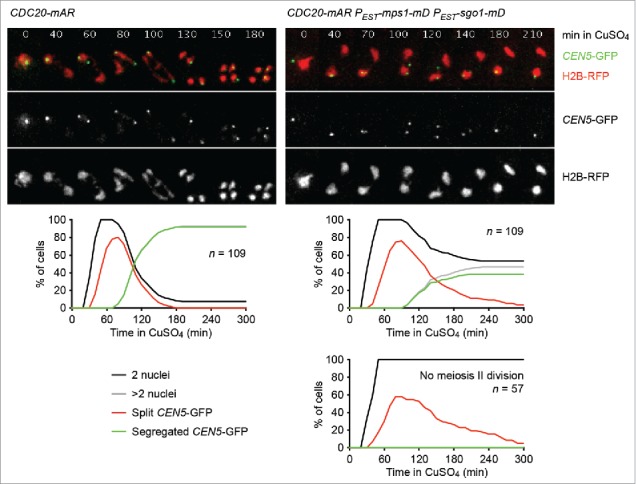
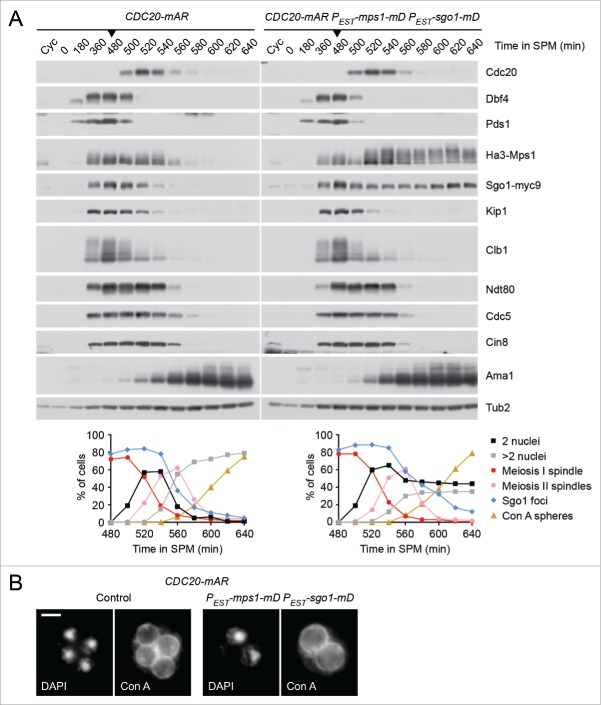
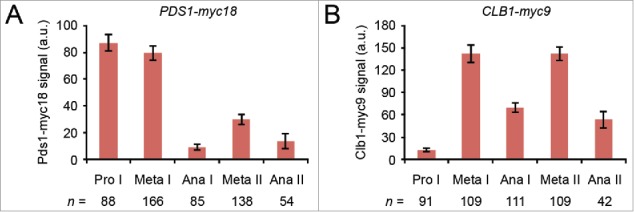
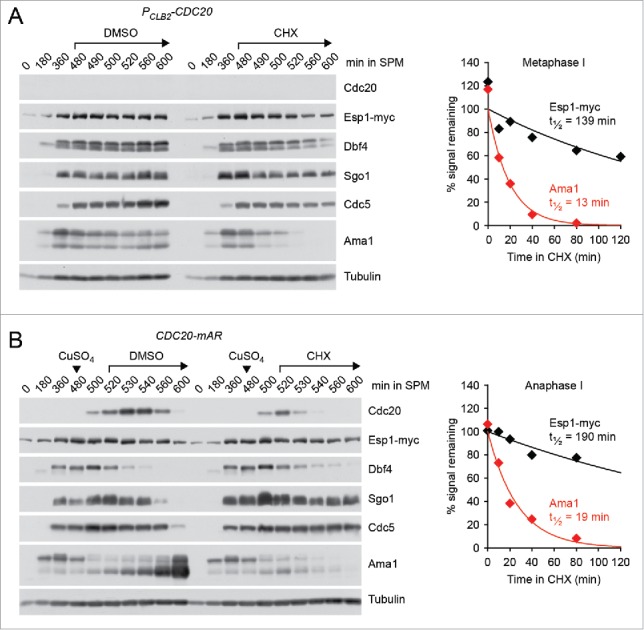
Cells undergoing meiosis produce haploid gametes through one round of DNA replication followed by 2 rounds of chromosome segregation. This requires that cohesin complexes, which establish sister chromatid cohesion during S phase, are removed in a stepwise manner. At meiosis I, the separase protease triggers the segregation of homologous chromosomes by cleaving cohesin's Rec8 subunit on chromosome arms. Cohesin persists at centromeres because the PP2A phosphatase, recruited by the shugoshin protein, dephosphorylates Rec8 and thereby protects it from cleavage. While chromatids disjoin upon cleavage of centromeric Rec8 at meiosis II, it was unclear how and when centromeric Rec8 is liberated from its protector PP2A. One proposal is that bipolar spindle forces separate PP2A from Rec8 as cells enter metaphase II. We show here that sister centromere biorientation is not sufficient to "deprotect" Rec8 at meiosis II in yeast. Instead, our data suggest that the ubiquitin-ligase APC/CCdc20 removes PP2A from centromeres by targeting for degradation the shugoshin Sgo1 and the kinase Mps1. This implies that Rec8 remains protected until entry into anaphase II when it is phosphorylated concurrently with the activation of separase. Here, we provide further support for this model and speculate on its relevance to mammalian oocytes.
Keywords: APC/C-Cdc20; Meiosis II; Mps1; PP2A; Rec8; centromere; cohesin cleavage; protection of centromeric cohesion; separase; shugoshin.
Figures
References
-
- Nasmyth K. Cohesin: a catenase with separate entry and exit gates? Nat Cell Biol 2011; 13:1170-7; PMID:21968990; https://doi.org/10.1038/ncb2349 - DOI - PubMed
-
- Haarhuis JHI, Elbatsh AMO, Rowland BD. Cohesin and Its Regulation: On the Logic of X-Shaped Chromosomes. Developmental Cell 2014; 31:7-18; PMID:25313959; https://doi.org/10.1016/j.devcel.2014.09.010 - DOI - PubMed
-
- Gligoris T, Lowe J. Structural Insights into Ring Formation of Cohesin and Related Smc Complexes. Trends Cell Biol 2016; 26:680-93; PMID:27134029; https://doi.org/10.1016/j.tcb.2016.04.002 - DOI - PMC - PubMed
-
- He X, Asthana S, Sorger PK. Transient sister chromatid separation and elastic deformation of chromosomes during mitosis in budding yeast. Cell 2000; 101:763-75; PMID:10892747; https://doi.org/10.1016/S0092-8674(00)80888-0 - DOI - PubMed
-
- Musacchio A. The Molecular Biology of Spindle Assembly Checkpoint Signaling Dynamics. Curr Biol 2015; 25:R1002-18. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases