

Influenza A Virus NS1 Protein Promotes Efficient Nuclear Export of Unspliced Viral M1 mRNA
- PMID: 28515301
- PMCID: PMC5651720
- DOI: 10.1128/JVI.00528-17
Influenza A Virus NS1 Protein Promotes Efficient Nuclear Export of Unspliced Viral M1 mRNA
Abstract
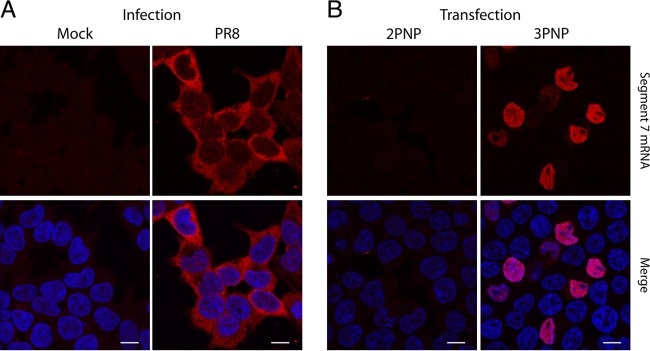
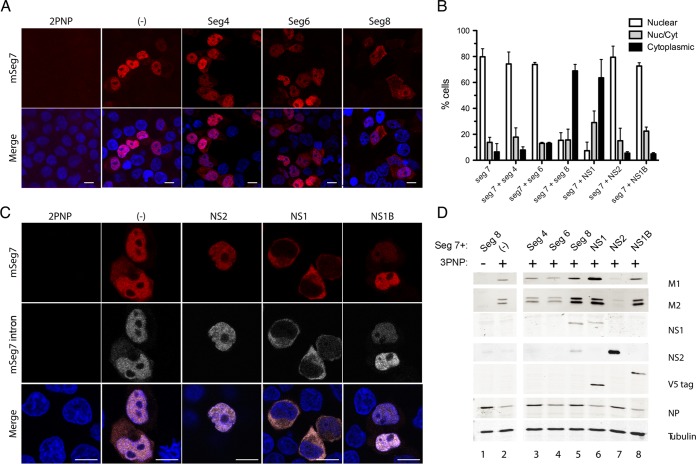
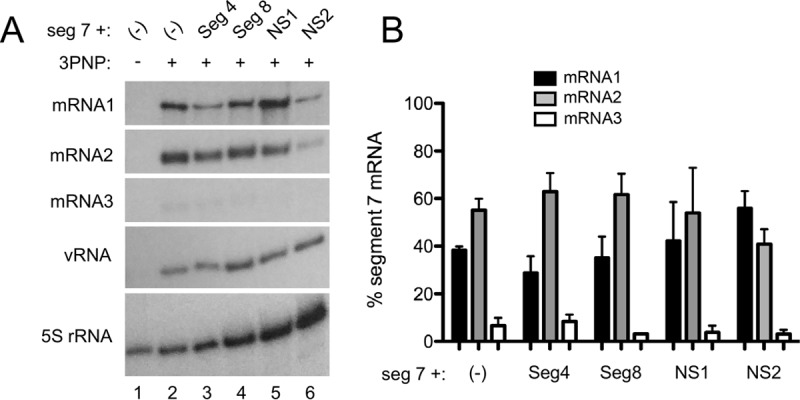
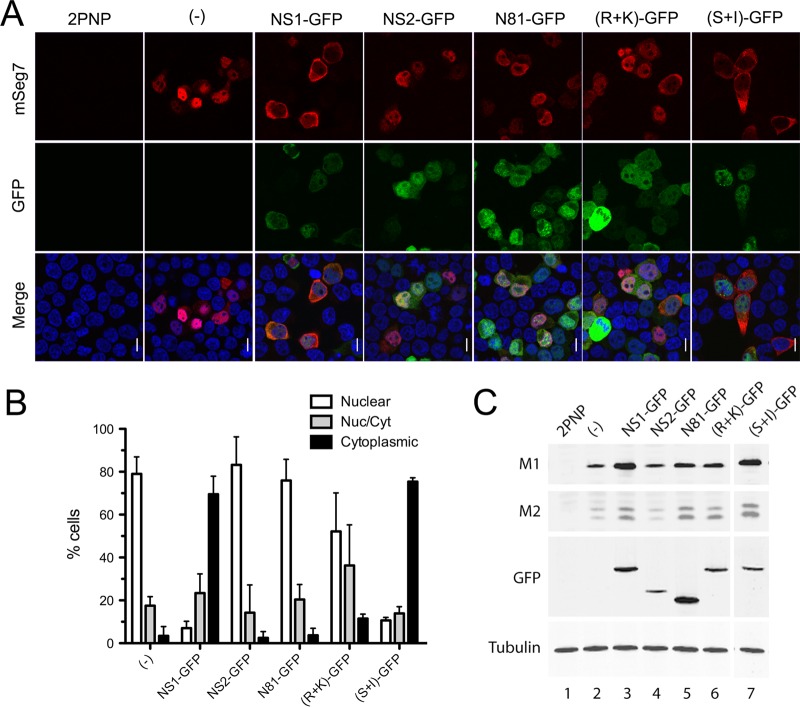
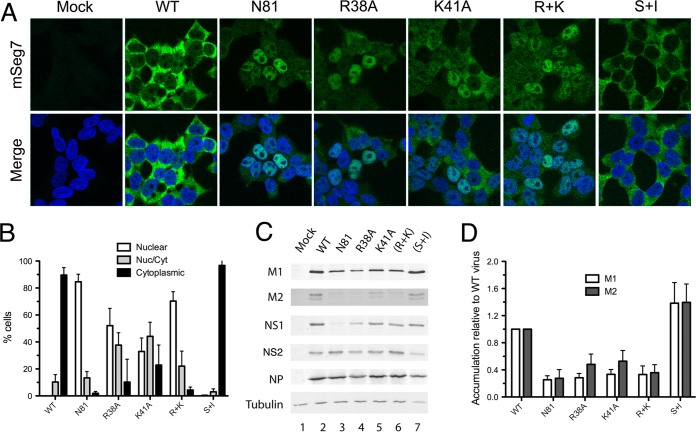
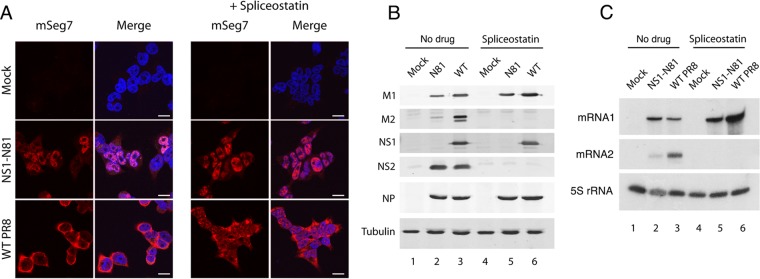
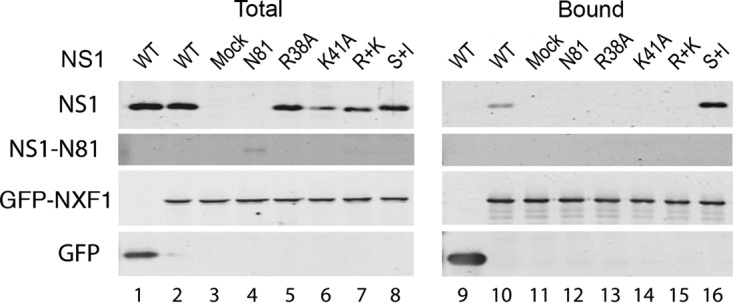
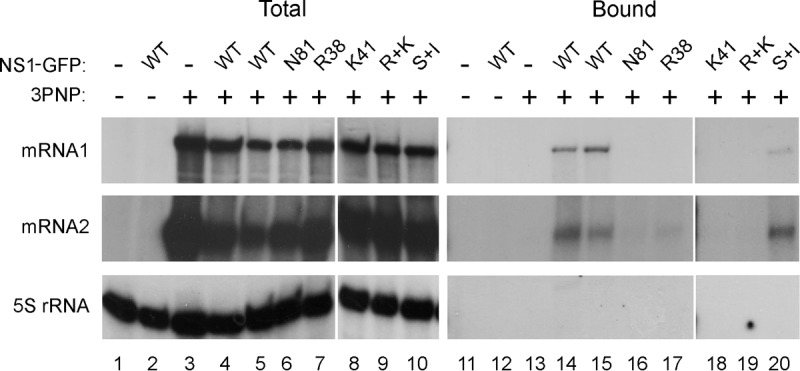
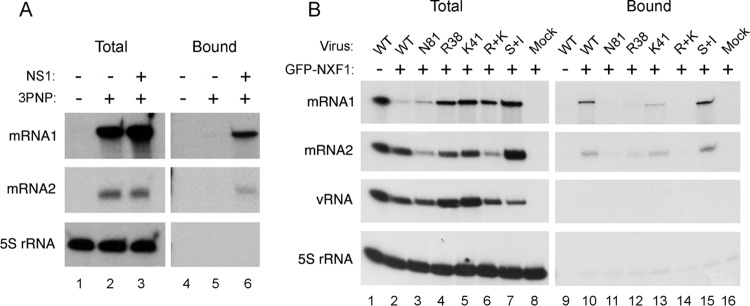
Influenza A virus mRNAs are transcribed by the viral RNA-dependent RNA polymerase in the cell nucleus before being exported to the cytoplasm for translation. Segment 7 produces two major transcripts: an unspliced mRNA that encodes the M1 matrix protein and a spliced transcript that encodes the M2 ion channel. Export of both mRNAs is dependent on the cellular NXF1/TAP pathway, but it is unclear how they are recruited to the export machinery or how the intron-containing but unspliced M1 mRNA bypasses the normal quality-control checkpoints. Using fluorescent in situ hybridization to monitor segment 7 mRNA localization, we found that cytoplasmic accumulation of unspliced M1 mRNA was inefficient in the absence of NS1, both in the context of segment 7 RNPs reconstituted by plasmid transfection and in mutant virus-infected cells. This effect was independent of any major effect on steady-state levels of segment 7 mRNA or splicing but corresponded to a ∼5-fold reduction in the accumulation of M1. A similar defect in intronless hemagglutinin (HA) mRNA nuclear export was seen with an NS1 mutant virus. Efficient export of M1 mRNA required both an intact NS1 RNA-binding domain and effector domain. Furthermore, while wild-type NS1 interacted with cellular NXF1 and also increased the interaction of segment 7 mRNA with NXF1, mutant NS1 polypeptides unable to promote mRNA export did neither. Thus, we propose that NS1 facilitates late viral gene expression by acting as an adaptor between viral mRNAs and the cellular nuclear export machinery to promote their nuclear export.IMPORTANCE Influenza A virus is a major pathogen of a wide variety of mammalian and avian species that threatens public health and food security. A fuller understanding of the virus life cycle is important to aid control strategies. The virus has a small genome that encodes relatively few proteins that are often multifunctional. Here, we characterize a new function for the NS1 protein, showing that, as well as previously identified roles in antagonizing the innate immune defenses of the cell and directly upregulating translation of viral mRNAs, it also promotes the nuclear export of the viral late gene mRNAs by acting as an adaptor between the viral mRNAs and the cellular mRNA nuclear export machinery.
Keywords: NS1; NXF1; influenza A virus mRNA export; nuclear import/export.
Copyright © 2017 Pereira et al.
Figures

References
-
- Inglis SC, Carroll AR, Lamb RA, Mahy BW. 1976. Polypeptides specified by the influenza virus genome. I. Evidence for eight distinct gene products specified by fowl plague virus. Virology 74:489–503. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
