

Control of Hsp90 chaperone and its clients by N-terminal acetylation and the N-end rule pathway
- PMID: 28515311
- PMCID: PMC5465900
- DOI: 10.1073/pnas.1705898114
Control of Hsp90 chaperone and its clients by N-terminal acetylation and the N-end rule pathway
Abstract
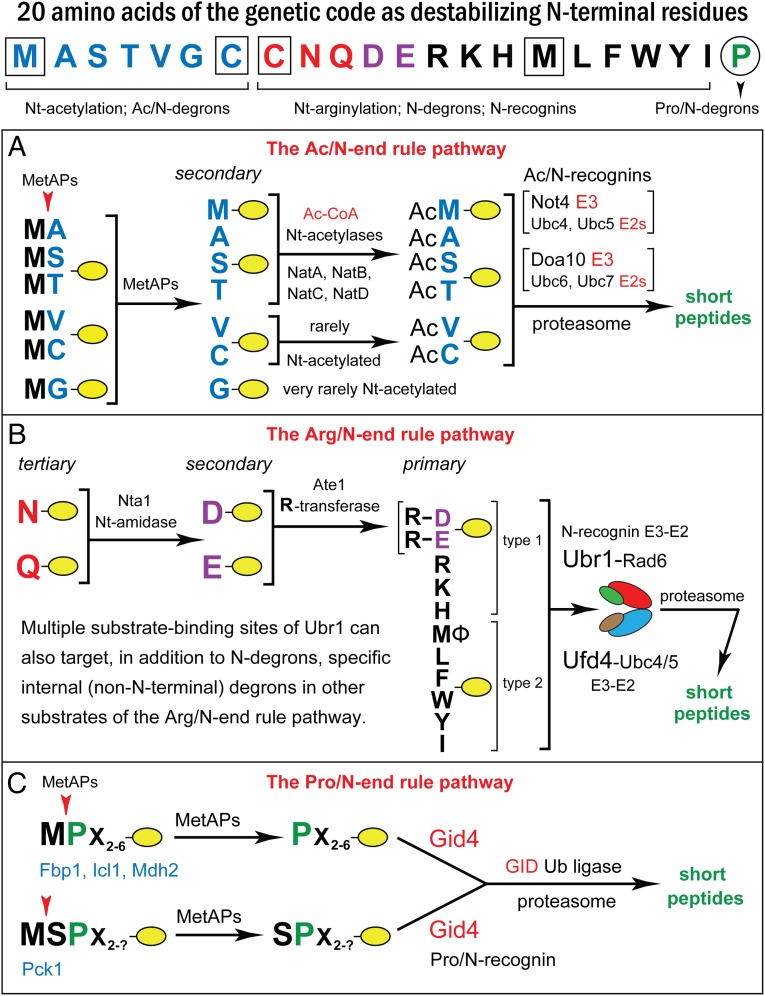
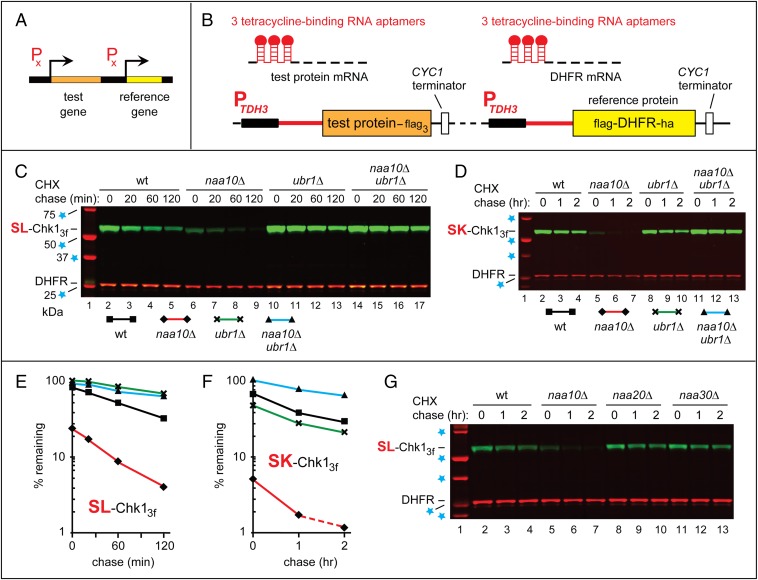
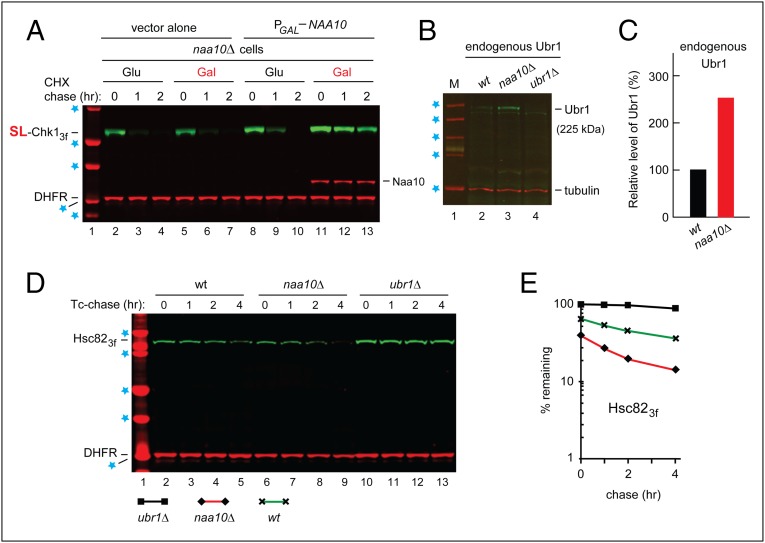
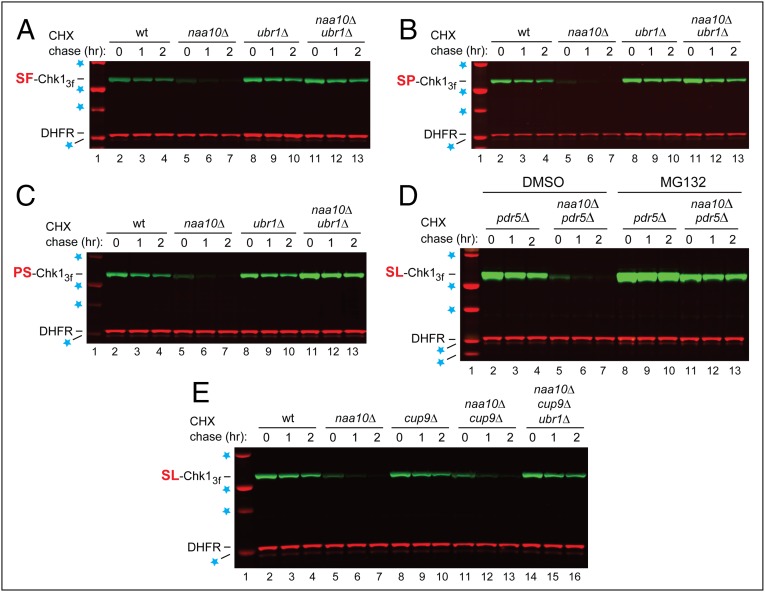
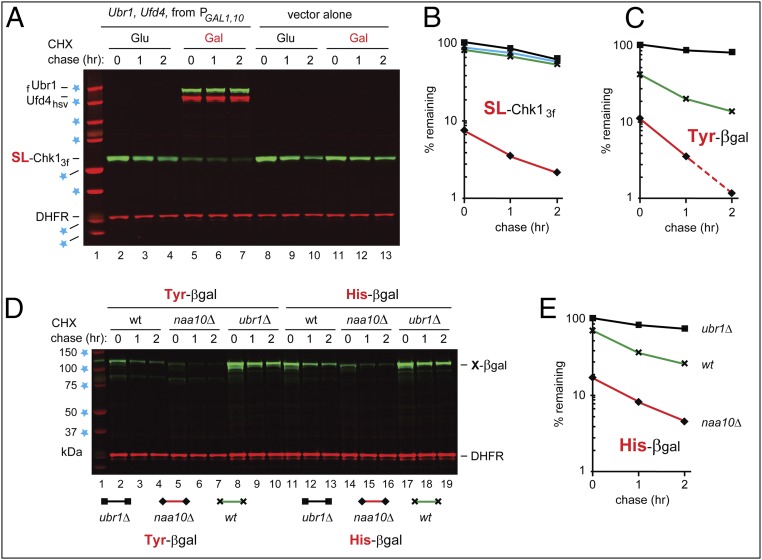
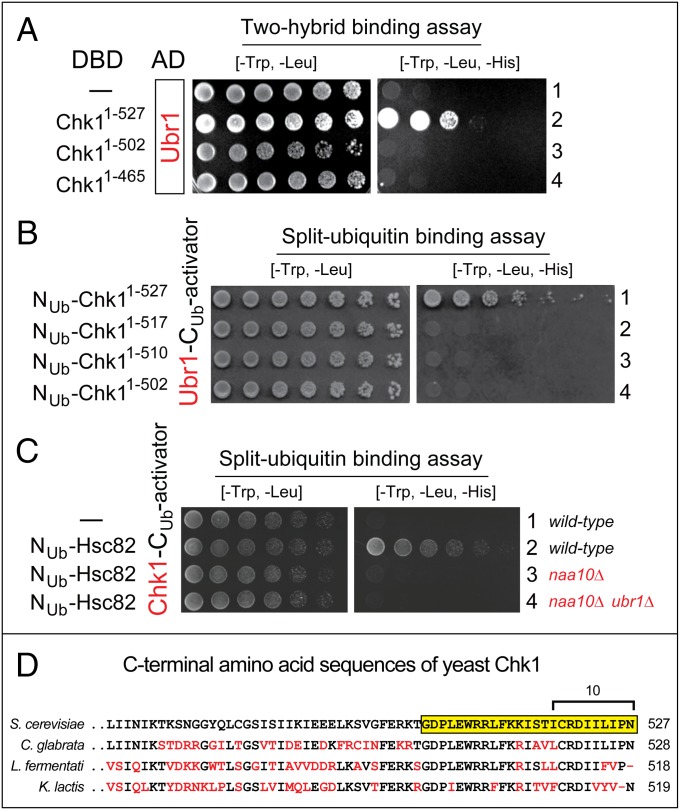
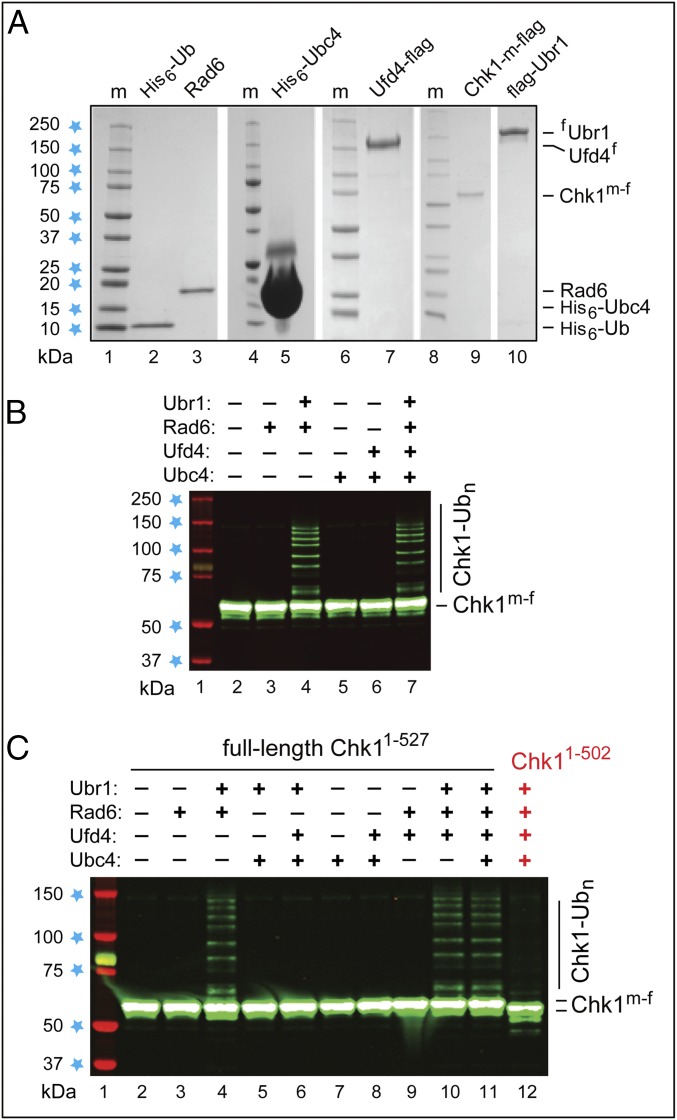
We found that the heat shock protein 90 (Hsp90) chaperone system of the yeast Saccharomyces cerevisiae is greatly impaired in naa10Δ cells, which lack the NatA Nα-terminal acetylase (Nt-acetylase) and therefore cannot N-terminally acetylate a majority of normally N-terminally acetylated proteins, including Hsp90 and most of its cochaperones. Chk1, a mitotic checkpoint kinase and a client of Hsp90, was degraded relatively slowly in wild-type cells but was rapidly destroyed in naa10Δ cells by the Arg/N-end rule pathway, which recognized a C terminus-proximal degron of Chk1. Diverse proteins (in addition to Chk1) that are shown here to be targeted for degradation by the Arg/N-end rule pathway in naa10Δ cells include Kar4, Tup1, Gpd1, Ste11, and also, remarkably, the main Hsp90 chaperone (Hsc82) itself. Protection of Chk1 by Hsp90 could be overridden not only by ablation of the NatA Nt-acetylase but also by overexpression of the Arg/N-end rule pathway in wild-type cells. Split ubiquitin-binding assays detected interactions between Hsp90 and Chk1 in wild-type cells but not in naa10Δ cells. These and related results revealed a major role of Nt-acetylation in the Hsp90-mediated protein homeostasis, a strong up-regulation of the Arg/N-end rule pathway in the absence of NatA, and showed that a number of Hsp90 clients are previously unknown substrates of the Arg/N-end rule pathway.
Keywords: Chk1; Naa10; Ubr1; Ufd4; protein degradation.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
The N-end rule pathway enzyme Naa10 supports epiblast specification in mouse embryonic stem cells by modulating FGF/MAPK.In Vitro Cell Dev Biol Anim. 2019 May;55(5):355-367. doi: 10.1007/s11626-019-00341-8. Epub 2019 Apr 16. In Vitro Cell Dev Biol Anim. 2019. PMID: 30993557
-
The N-end rule pathway is mediated by a complex of the RING-type Ubr1 and HECT-type Ufd4 ubiquitin ligases.Nat Cell Biol. 2010 Dec;12(12):1177-85. doi: 10.1038/ncb2121. Epub 2010 Nov 14. Nat Cell Biol. 2010. PMID: 21076411 Free PMC article.
-
A Saccharomyces cerevisiae model reveals in vivo functional impairment of the Ogden syndrome N-terminal acetyltransferase NAA10 Ser37Pro mutant.Mol Cell Proteomics. 2014 Aug;13(8):2031-41. doi: 10.1074/mcp.M113.035402. Epub 2014 Jan 9. Mol Cell Proteomics. 2014. PMID: 24408909 Free PMC article.
-
The N-end rule pathway and regulation by proteolysis.Protein Sci. 2011 Aug;20(8):1298-345. doi: 10.1002/pro.666. Protein Sci. 2011. PMID: 21633985 Free PMC article. Review.
-
Molecular interaction network of the Hsp90 chaperone system.Adv Exp Med Biol. 2007;594:27-36. doi: 10.1007/978-0-387-39975-1_3. Adv Exp Med Biol. 2007. PMID: 17205672 Review.
Cited by
-
Gid10 as an alternative N-recognin of the Pro/N-degron pathway.Proc Natl Acad Sci U S A. 2019 Aug 6;116(32):15914-15923. doi: 10.1073/pnas.1908304116. Epub 2019 Jul 23. Proc Natl Acad Sci U S A. 2019. PMID: 31337681 Free PMC article.
-
Co-translational, Post-translational, and Non-catalytic Roles of N-Terminal Acetyltransferases.Mol Cell. 2019 Mar 21;73(6):1097-1114. doi: 10.1016/j.molcel.2019.02.007. Epub 2019 Mar 13. Mol Cell. 2019. PMID: 30878283 Free PMC article. Review.
-
Evolution of Substrates and Components of the Pro/N-Degron Pathway.Biochemistry. 2020 Feb 4;59(4):582-593. doi: 10.1021/acs.biochem.9b00953. Epub 2020 Jan 2. Biochemistry. 2020. PMID: 31895557 Free PMC article.
-
Control of protein degradation by N-terminal acetylation and the N-end rule pathway.Exp Mol Med. 2018 Jul 27;50(7):1-8. doi: 10.1038/s12276-018-0097-y. Exp Mol Med. 2018. PMID: 30054456 Free PMC article. Review.
-
N-degron and C-degron pathways of protein degradation.Proc Natl Acad Sci U S A. 2019 Jan 8;116(2):358-366. doi: 10.1073/pnas.1816596116. Proc Natl Acad Sci U S A. 2019. PMID: 30622213 Free PMC article. Review.
References
-
- Hartl FU, Bracher A, Hayer-Hartl M. Molecular chaperones in protein folding and homeostasis. Nature. 2011;475:324–332. - PubMed
-
- Horwich AL. Molecular chaperones in cellular protein folding: The birth of a field. Cell. 2014;157:285–288. - PubMed
-
- Wolff S, Weissman JS, Dillin A. Differential scales of protein quality control. Cell. 2014;157:52–64. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
