

Assessment of trophic ecomorphology in non-alligatoroid crocodylians and its adaptive and taxonomic implications
- PMID: 28516735
- PMCID: PMC5522899
- DOI: 10.1111/joa.12626
Assessment of trophic ecomorphology in non-alligatoroid crocodylians and its adaptive and taxonomic implications
Abstract
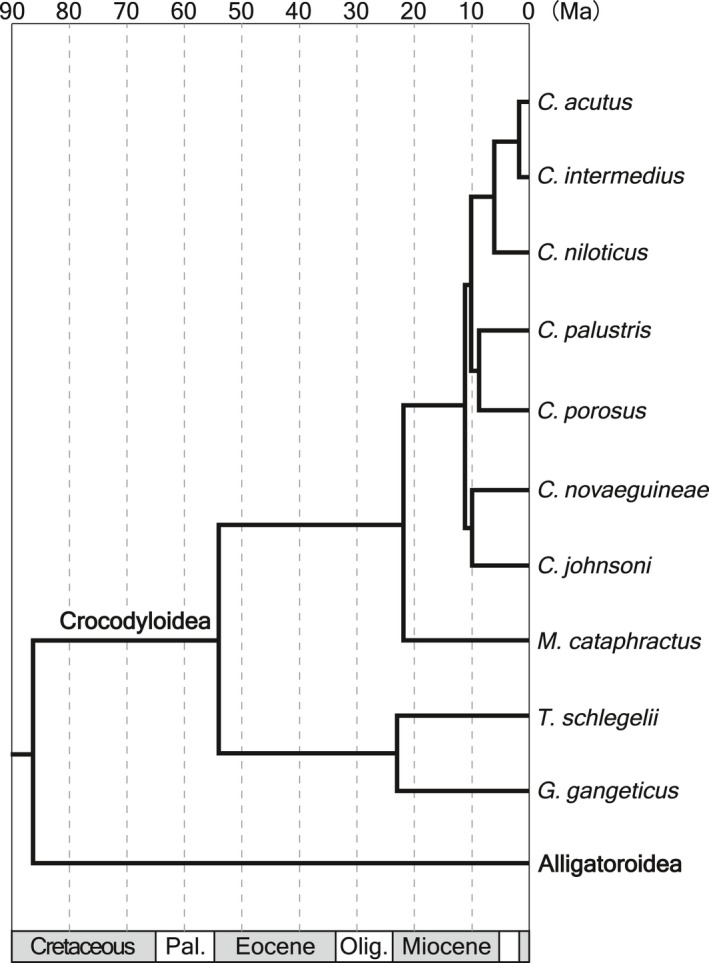
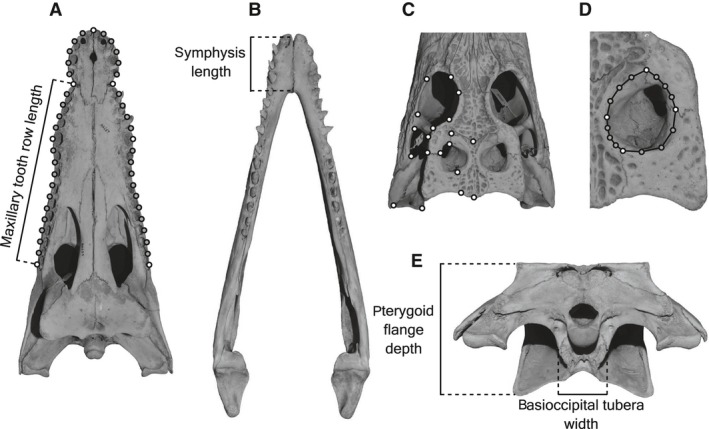
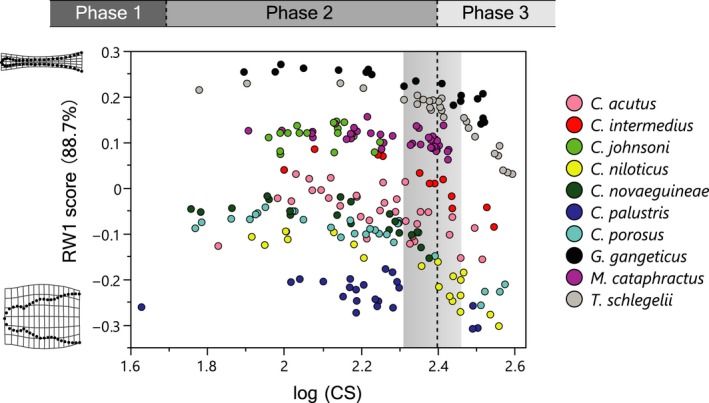
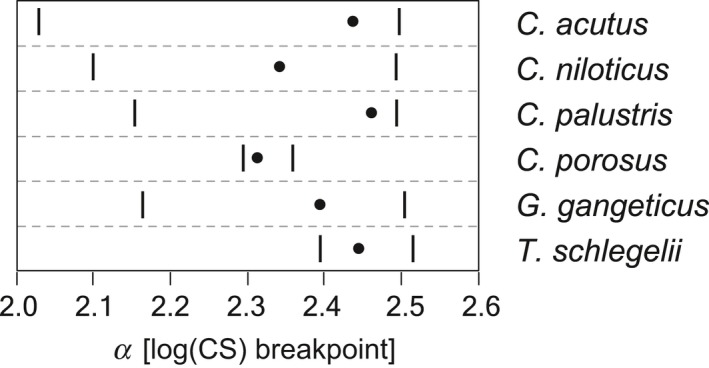
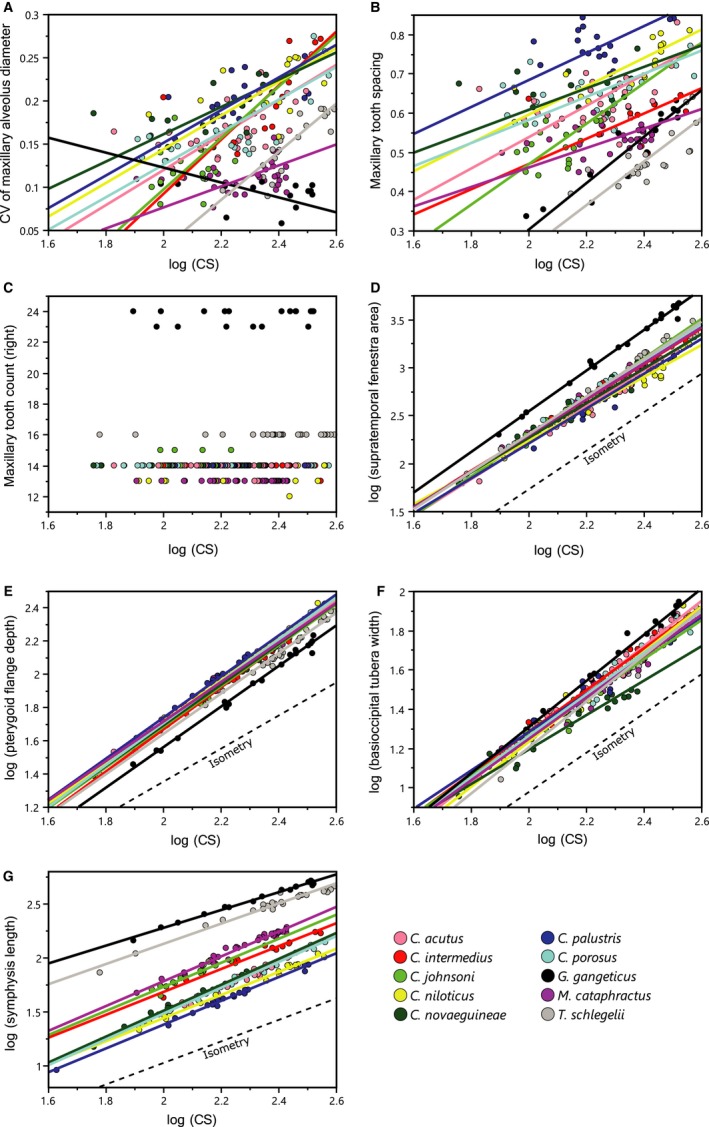
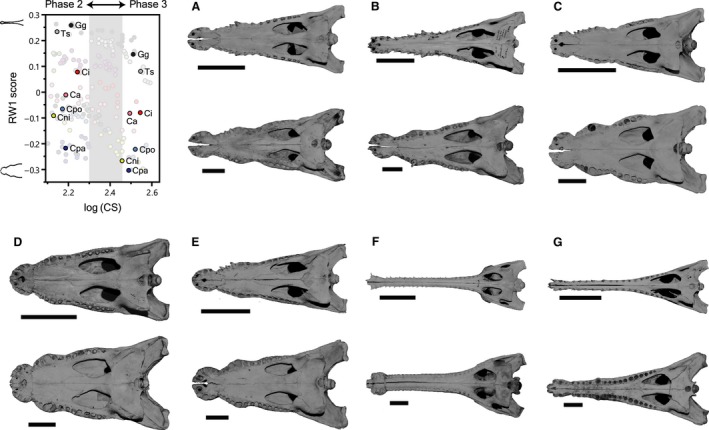
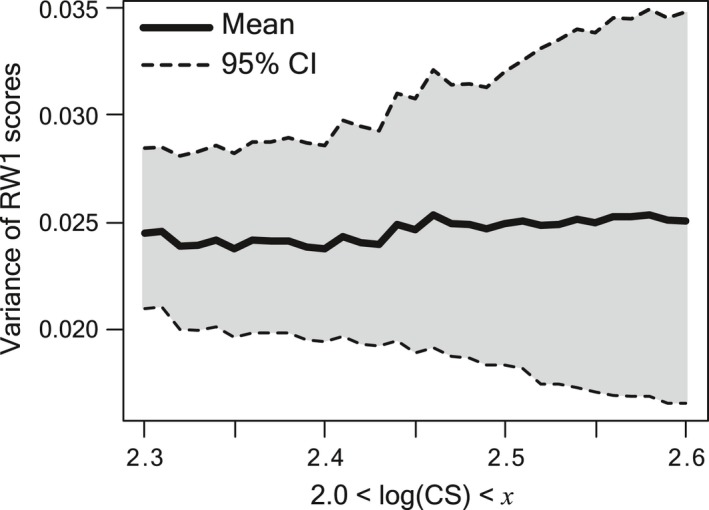
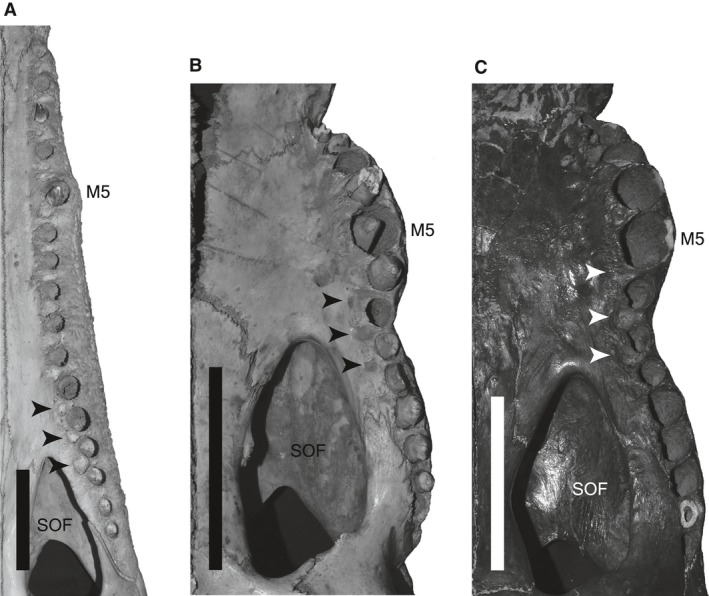
Although the establishment of trophic ecomorphology in living crocodylians can contribute to estimating feeding habits of extinct large aquatic reptiles, assessment of ecomorphological traits other than the snout shape has scarcely been conducted in crocodylians. Here, I tested the validity of the proposed trophic ecomorphological traits in crocodylians by examining the correlation between those traits and the snout shape (an established trophic ecomorphology), using 10 non-alligatoroid crocodylian species with a wide range of snout shape. I then compared the ontogenetic scaling of trophic ecomorphology to discuss its adaptive and taxonomic significance. The results demonstrated that degree of heterodonty, tooth spacing, size of supratemporal fenestra (STF), ventral extension of pterygoid flange and length of lower jaw symphysis are significantly correlated with snout shape by both non-phylogenetic and phylogenetic regression analyses. Gavialis gangeticus falls outside of 95% prediction intervals for the relationships of some traits and the snout shape, suggesting that piscivorous specialization involves the deviation from the typical transformation axis of skull characters. The comparative snout shape ontogeny revealed a universal trend of snout widening through growth in the sampled crocodylians, implying the existence of a shared size-dependent biomechanical constraint in non-alligatoroid crocodylians. Growth patterns of other traits indicated that G. gangeticus shows atypical trends for degree of heterodonty, size of STF, and symphysis length, whereas the same trends are shared for tooth spacing and ventral extension of pterygoid flange among non-alligatoroid crocodylians. These suggest that some characters are ontogenetically labile in response to prey preference shifts through growth, but other characters are in keeping with the conserved biomechanics among non-alligatoroid crocodylians. Some important taxonomic characters such as the occlusal pattern are likely correlated with ontogeny and trophic ecomorphology rather than are constrained by phylogenetic relationships, and careful reassessment of such characters might be necessary for better reconstructing the morphological phylogeny of crocodylians.
Keywords: allometry; crocodylian; occlusal pattern; piscivory; trophic ecomorphology.
© 2017 Anatomical Society.
Figures




References
-
- Arnold SJ (1983) Morphology, performance and fitness. Am Zool 23, 347–361.
-
- Bock WJ (1990) From biologische anatomie to ecomorphology. Netherl J Zool 40, 254–277.
-
- Bona P, Desojo JB (2011) Osteology and cranial musculature of Caiman latirostris (Crocodylia: Alligatoridae). J Morphol 272, 780–795. - PubMed
-
- Bookstein FL (1991) Morphometric Tools for Landmark Data. Cambridge: Cambridge University Press.
-
- Brochu CA (1997) Morphology, fossils, divergence timing, and the phylogenetic relationships of Gavialis . Syst Biol 46, 479–522. - PubMed
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
