

Maternal age-dependent APC/C-mediated decrease in securin causes premature sister chromatid separation in meiosis II
- PMID: 28516917
- PMCID: PMC5454377
- DOI: 10.1038/ncomms15346
Maternal age-dependent APC/C-mediated decrease in securin causes premature sister chromatid separation in meiosis II
Abstract
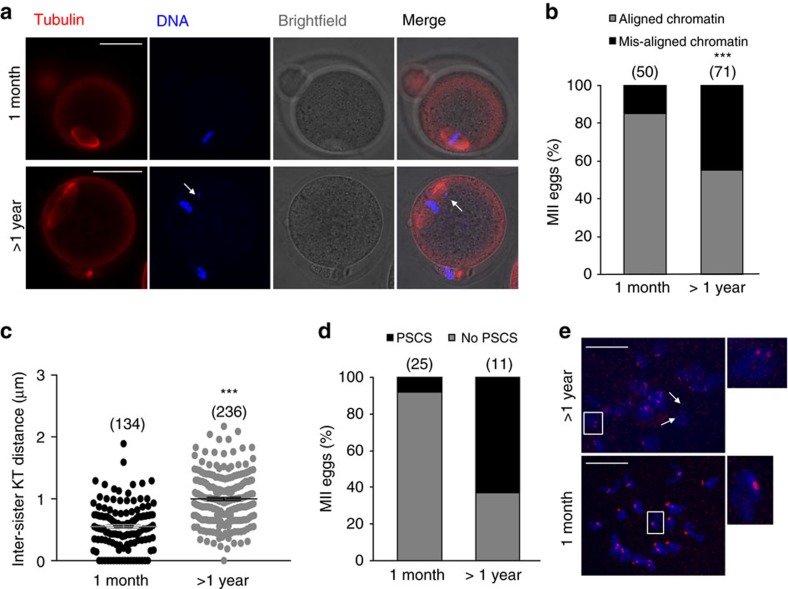
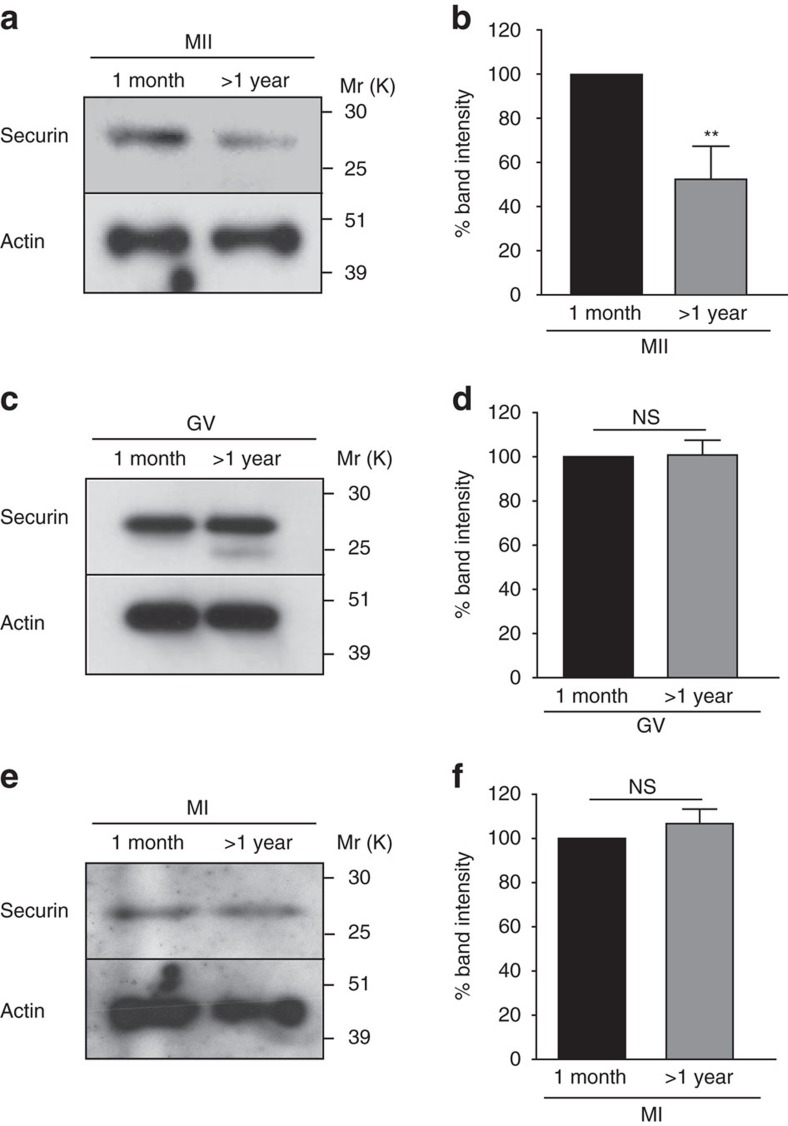
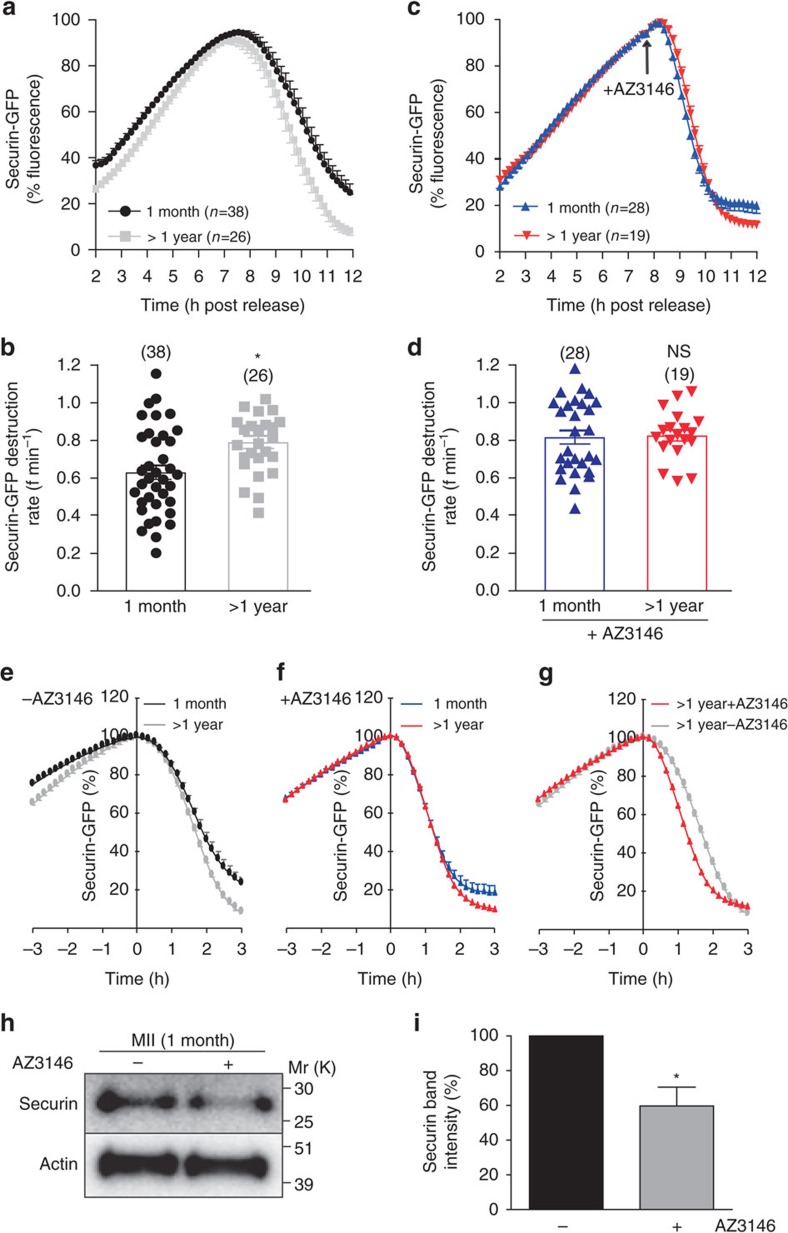
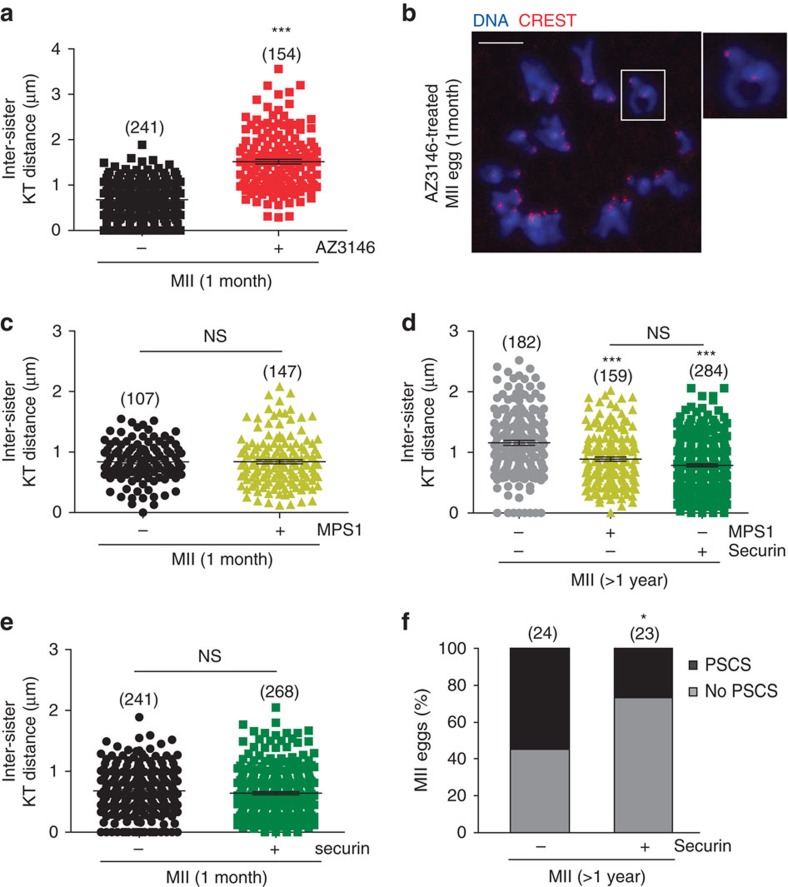
Sister chromatid attachment during meiosis II (MII) is maintained by securin-mediated inhibition of separase. In maternal ageing, oocytes show increased inter-sister kinetochore distance and premature sister chromatid separation (PSCS), suggesting aberrant separase activity. Here, we find that MII oocytes from aged mice have less securin than oocytes from young mice and that this reduction is mediated by increased destruction by the anaphase promoting complex/cyclosome (APC/C) during meiosis I (MI) exit. Inhibition of the spindle assembly checkpoint (SAC) kinase, Mps1, during MI exit in young oocytes replicates this phenotype. Further, over-expression of securin or Mps1 protects against the age-related increase in inter-sister kinetochore distance and PSCS. These findings show that maternal ageing compromises the oocyte SAC-APC/C axis leading to a decrease in securin that ultimately causes sister chromatid cohesion loss. Manipulating this axis and/or increasing securin may provide novel therapeutic approaches to alleviating the risk of oocyte aneuploidy in maternal ageing.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
Similar articles
-
Securin regulates the spatiotemporal dynamics of separase.J Cell Biol. 2025 Feb 3;224(2):e202312099. doi: 10.1083/jcb.202312099. Epub 2024 Nov 18. J Cell Biol. 2025. PMID: 39556062 Free PMC article.
-
CENP-F-dependent DRP1 function regulates APC/C activity during oocyte meiosis I.Nat Commun. 2022 Dec 13;13(1):7732. doi: 10.1038/s41467-022-35461-5. Nat Commun. 2022. PMID: 36513638 Free PMC article.
-
Spindle formation, chromosome segregation and the spindle checkpoint in mammalian oocytes and susceptibility to meiotic error.Mutat Res. 2008 Mar 12;651(1-2):14-29. doi: 10.1016/j.mrgentox.2007.10.015. Epub 2007 Nov 9. Mutat Res. 2008. PMID: 18096427 Review.
-
Regulation of APC/C activity in oocytes by a Bub1-dependent spindle assembly checkpoint.Curr Biol. 2009 Mar 10;19(5):369-80. doi: 10.1016/j.cub.2009.01.064. Epub 2009 Feb 26. Curr Biol. 2009. PMID: 19249208
-
Separase and Roads to Disengage Sister Chromatids during Anaphase.Int J Mol Sci. 2023 Feb 27;24(5):4604. doi: 10.3390/ijms24054604. Int J Mol Sci. 2023. PMID: 36902034 Free PMC article. Review.
Cited by
-
UBE2V1 governs aging induced protein aggregation and developmental defects in oocytes and embryos.Commun Biol. 2025 May 21;8(1):769. doi: 10.1038/s42003-025-08214-5. Commun Biol. 2025. PMID: 40394227 Free PMC article.
-
Ribonucleic Acid Export 1 Is a Kinetochore-Associated Protein That Participates in Chromosome Alignment in Mouse Oocytes.Int J Mol Sci. 2021 May 3;22(9):4841. doi: 10.3390/ijms22094841. Int J Mol Sci. 2021. PMID: 34063622 Free PMC article.
-
Multi-omics reveal the metabolic patterns in mouse cumulus cells during oocyte maturation.J Ovarian Res. 2023 Aug 8;16(1):156. doi: 10.1186/s13048-023-01237-8. J Ovarian Res. 2023. PMID: 37550748 Free PMC article.
-
Cell Division Cycle 5-Like Regulates Metaphase-to-Anaphase Transition in Meiotic Oocyte.Front Cell Dev Biol. 2021 Jul 1;9:671685. doi: 10.3389/fcell.2021.671685. eCollection 2021. Front Cell Dev Biol. 2021. PMID: 34277613 Free PMC article.
-
Hallmarks of female reproductive aging in physiologic aging mice.Nat Aging. 2024 Dec;4(12):1711-1730. doi: 10.1038/s43587-024-00769-y. Epub 2024 Dec 13. Nat Aging. 2024. PMID: 39672896 Free PMC article. Review.
References
-
- Hassold T. & Hunt P. To ERR (meiotically) is human: the genesis of human aneuploidy. Nat. Rev. Genet. 2, 280–291 (2001). - PubMed
-
- Hassold T., Hall H. & Hunt P. The origin of human aneuploidy: where we have been, where we are going. Hum. Mol. Genet. 16, R203–R208 (2007). - PubMed
-
- Angell R. R. Predivision in human oocytes at meiosis-I-A mechanism for trisomy formation in man. Hum. Genet. 86, 383–387 (1991). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
