

Translational control and Rho-dependent transcription termination are intimately linked in riboswitch regulation
- PMID: 28520932
- PMCID: PMC5499598
- DOI: 10.1093/nar/gkx434
Translational control and Rho-dependent transcription termination are intimately linked in riboswitch regulation
Abstract
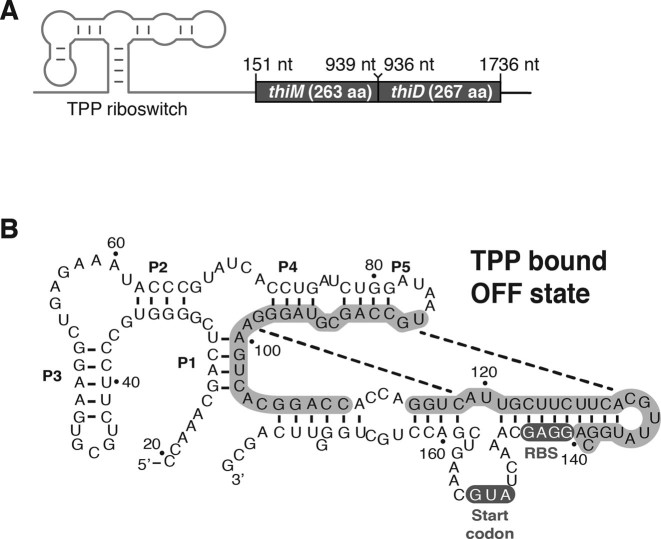
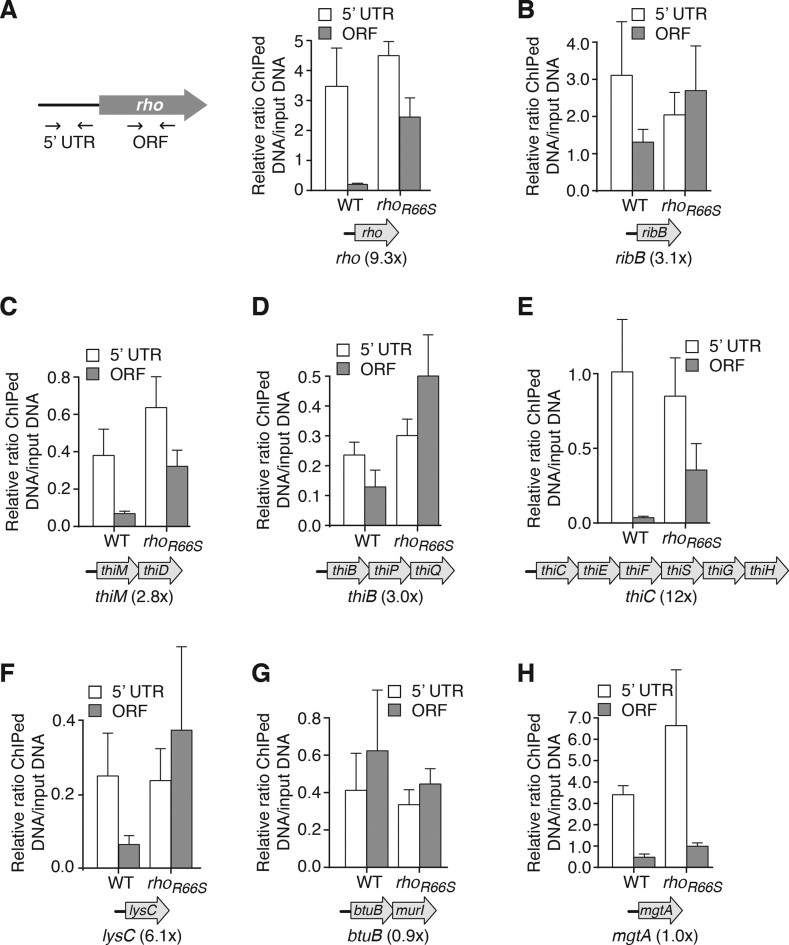
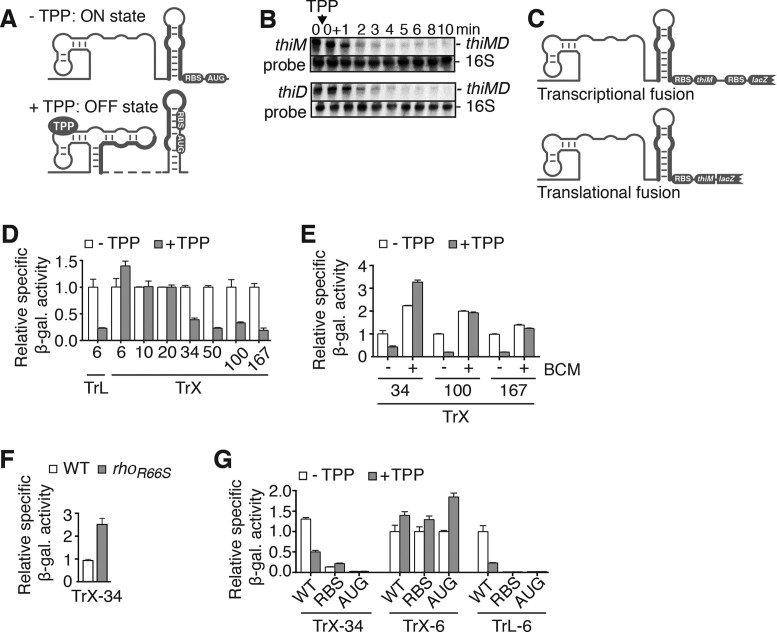
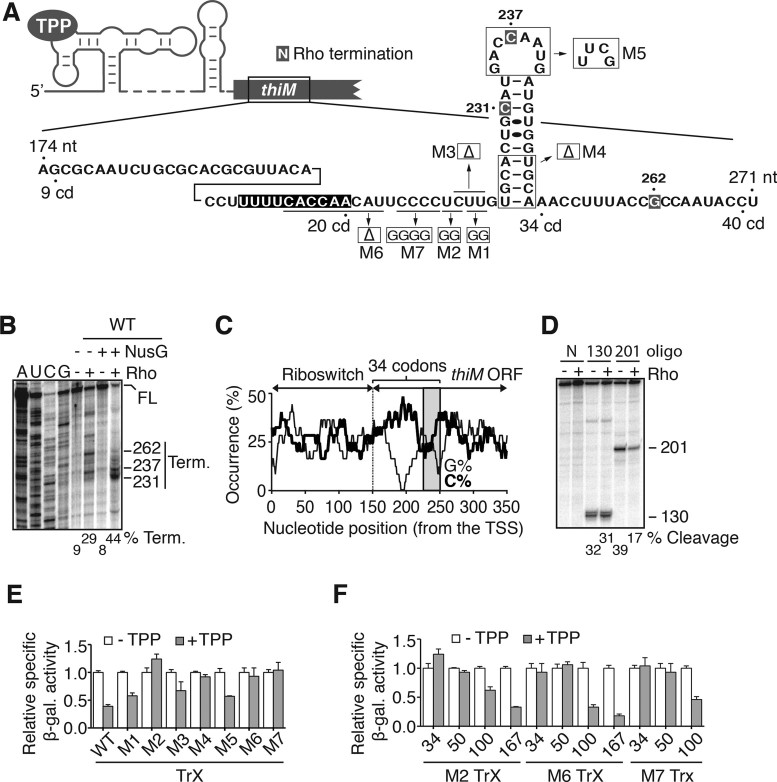
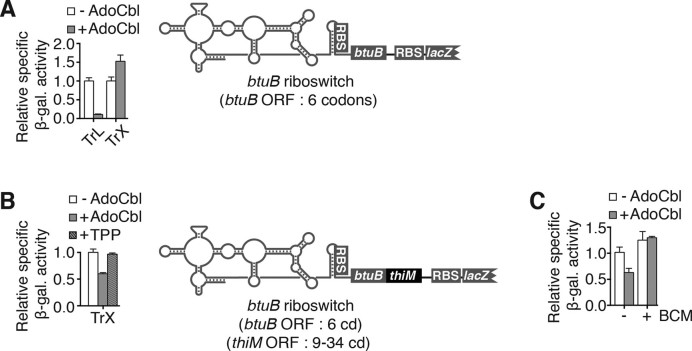
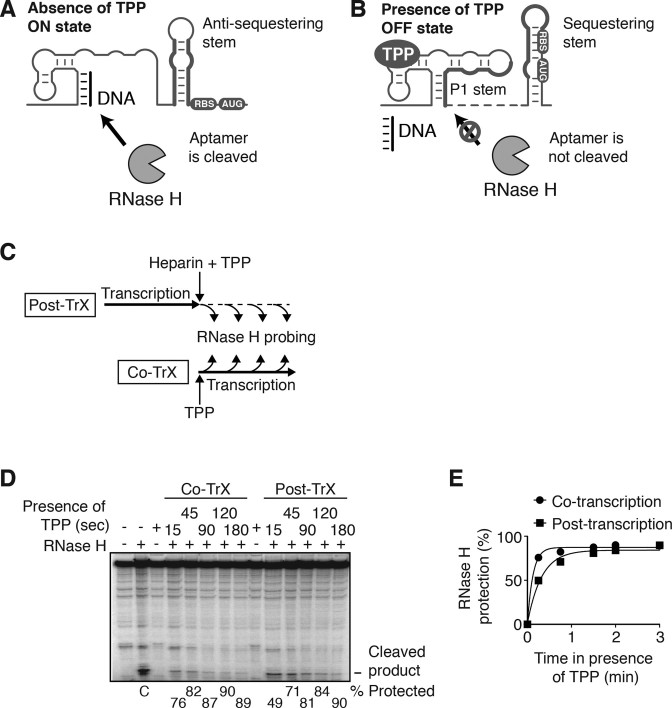
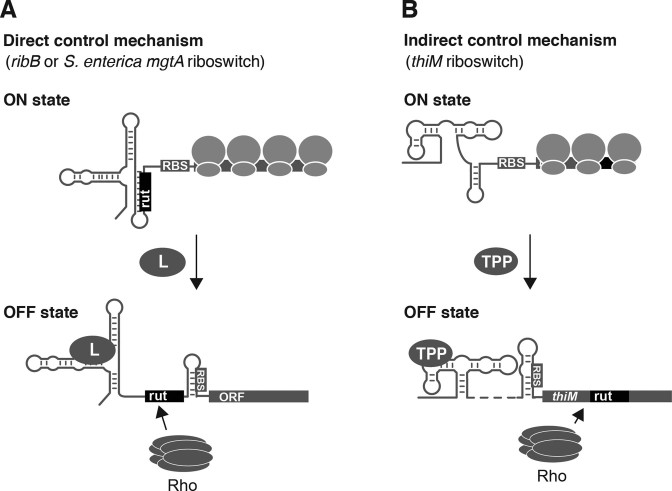
Riboswitches are regulatory elements that control gene expression by altering RNA structure upon the binding of specific metabolites. Although Bacillus subtilis riboswitches have been shown to control premature transcription termination, less is known about regulatory mechanisms employed by Escherichia coli riboswitches, which are predicted to regulate mostly at the level of translation initiation. Here, we present experimental evidence suggesting that the majority of known E. coli riboswitches control transcription termination by using the Rho transcription factor. In the case of the thiamin pyrophosphate-dependent thiM riboswitch, we find that Rho-dependent transcription termination is triggered as a consequence of translation repression. Using in vitro and in vivo assays, we show that the Rho-mediated regulation relies on RNA target elements located at the beginning of thiM coding region. Gene reporter assays indicate that relocating Rho target elements to a different gene induces transcription termination, demonstrating that such elements are modular domains controlling Rho. Our work provides strong evidence that translationally regulating riboswitches also regulate mRNA levels through an indirect control mechanism ensuring tight control of gene expression.
© The Author(s) 2017. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures
References
-
- Haller A., Souliere M.F., Micura R.. The dynamic nature of RNA as key to understanding riboswitch mechanisms. Acc. Chem. Res. 2011; 44:1339–1348. - PubMed
-
- Loh E., Dussurget O., Gripenland J., Vaitkevicius K., Tiensuu T., Mandin P., Repoila F., Buchrieser C., Cossart P., Johansson J.. A trans-acting riboswitch controls expression of the virulence regulator PrfA in Listeria monocytogenes. Cell. 2009; 139:770–779. - PubMed
-
- Mellin J.R., Koutero M., Dar D., Nahori M.-A., Sorek R., Cossart P.. Sequestration of a two-component response regulator by a riboswitch-regulated noncoding RNA. Science. 2014; 345:940–943. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
