

The DEAD-box helicase Mss116 plays distinct roles in mitochondrial ribogenesis and mRNA-specific translation
- PMID: 28520979
- PMCID: PMC5499750
- DOI: 10.1093/nar/gkx426
The DEAD-box helicase Mss116 plays distinct roles in mitochondrial ribogenesis and mRNA-specific translation
Abstract
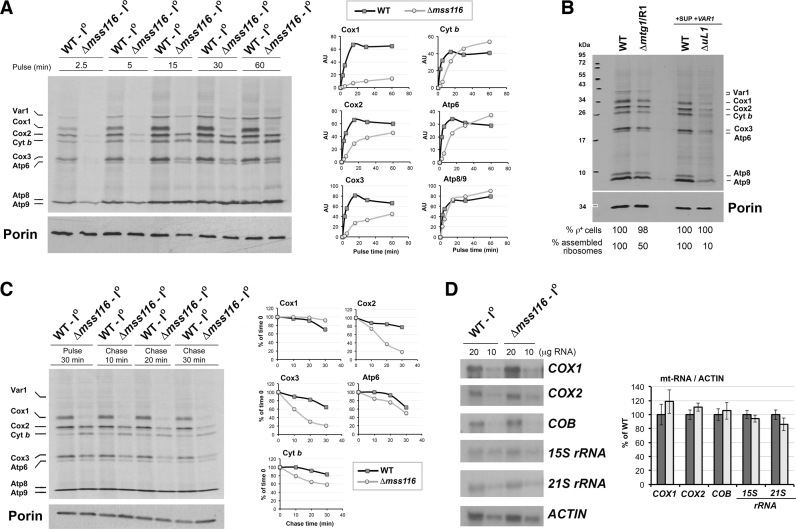
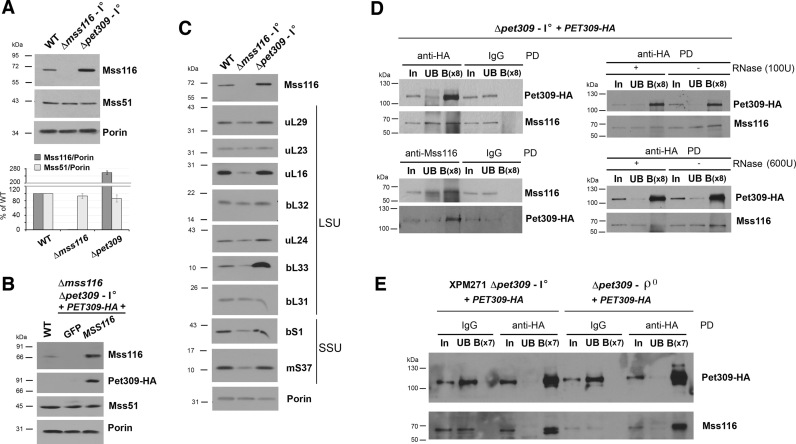
Members of the DEAD-box family are often multifunctional proteins involved in several RNA transactions. Among them, yeast Saccharomyces cerevisiae Mss116 participates in mitochondrial intron splicing and, under cold stress, also in mitochondrial transcription elongation. Here, we show that Mss116 interacts with the mitoribosome assembly factor Mrh4, is required for efficient mitoribosome biogenesis, and consequently, maintenance of the overall mitochondrial protein synthesis rate. Additionally, Mss116 is required for efficient COX1 mRNA translation initiation and elongation. Mss116 interacts with a COX1 mRNA-specific translational activator, the pentatricopeptide repeat protein Pet309. In the absence of Mss116, Pet309 is virtually absent, and although mitoribosome loading onto COX1 mRNA can occur, activation of COX1 mRNA translation is impaired. Mutations abolishing the helicase activity of Mss116 do not prevent the interaction of Mss116 with Pet309 but also do not allow COX1 mRNA translation. We propose that Pet309 acts as an adaptor protein for Mss116 action on the COX1 mRNA 5΄-UTR to promote efficient Cox1 synthesis. Overall, we conclude that the different functions of Mss116 in the biogenesis and functioning of the mitochondrial translation machinery depend on Mss116 interplay with its protein cofactors.
© The Author(s) 2017. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures






Similar articles
-
The Pet309 pentatricopeptide repeat motifs mediate efficient binding to the mitochondrial COX1 transcript in yeast.RNA Biol. 2014;11(7):953-67. doi: 10.4161/rna.29780. Epub 2014 Jul 24. RNA Biol. 2014. PMID: 25181249 Free PMC article.
-
Insights into the translational activation mechanisms of the COX1 mRNA in yeast mitochondria.J Cell Sci. 2025 Aug 15;138(16):jcs263694. doi: 10.1242/jcs.263694. Epub 2025 Sep 2. J Cell Sci. 2025. PMID: 40891829
-
The product of the nuclear gene PET309 is required for translation of mature mRNA and stability or production of intron-containing RNAs derived from the mitochondrial COX1 locus of Saccharomyces cerevisiae.EMBO J. 1995 Aug 15;14(16):4031-43. doi: 10.1002/j.1460-2075.1995.tb00074.x. EMBO J. 1995. PMID: 7664742 Free PMC article.
-
Yeast and human mitochondrial helicases.Biochim Biophys Acta. 2013 Aug;1829(8):842-53. doi: 10.1016/j.bbagrm.2013.02.009. Epub 2013 Feb 27. Biochim Biophys Acta. 2013. PMID: 23454114 Review.
-
Cytochrome c oxidase biogenesis - from translation to early assembly of the core subunit COX1.FEBS Lett. 2023 Jun;597(12):1569-1578. doi: 10.1002/1873-3468.14671. Epub 2023 May 31. FEBS Lett. 2023. PMID: 37247261 Review.
Cited by
-
Yeast Mitoribosome Large Subunit Assembly Proceeds by Hierarchical Incorporation of Protein Clusters and Modules on the Inner Membrane.Cell Metab. 2018 Mar 6;27(3):645-656.e7. doi: 10.1016/j.cmet.2018.01.012. Cell Metab. 2018. PMID: 29514071 Free PMC article.
-
The translational activator Sov1 coordinates mitochondrial gene expression with mitoribosome biogenesis.Nucleic Acids Res. 2020 Jul 9;48(12):6759-6774. doi: 10.1093/nar/gkaa424. Nucleic Acids Res. 2020. PMID: 32449921 Free PMC article.
-
IRC3 Regulates Mitochondrial Translation in Response to Metabolic Cues in Saccharomyces cerevisiae.Mol Cell Biol. 2021 Oct 26;41(11):e0023321. doi: 10.1128/MCB.00233-21. Epub 2021 Aug 16. Mol Cell Biol. 2021. PMID: 34398681 Free PMC article.
-
Mitochondrial ribosome bL34 mutants present diminished translation of cytochrome c oxidase subunits.Cell Biol Int. 2018 Jun;42(6):630-642. doi: 10.1002/cbin.10913. Epub 2017 Dec 7. Cell Biol Int. 2018. PMID: 29160602 Free PMC article.
-
Cardiomyopathy-associated mutation in the ADP/ATP carrier reveals translation-dependent regulation of cytochrome c oxidase activity.Mol Biol Cell. 2018 Jun 15;29(12):1449-1464. doi: 10.1091/mbc.E17-12-0700. Epub 2018 Apr 24. Mol Biol Cell. 2018. PMID: 29688796 Free PMC article.
References
-
- Greber B.J., Bieri P., Leibundgut M., Leitner A., Aebersold R., Boehringer D., Ban N.. Ribosome. The complete structure of the 55S mammalian mitochondrial ribosome. Science. 2015; 348:303–308. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
