

Long terminal repeats power evolution of genes and gene expression programs in mammalian oocytes and zygotes
- PMID: 28522611
- PMCID: PMC5538554
- DOI: 10.1101/gr.216150.116
Long terminal repeats power evolution of genes and gene expression programs in mammalian oocytes and zygotes
Abstract
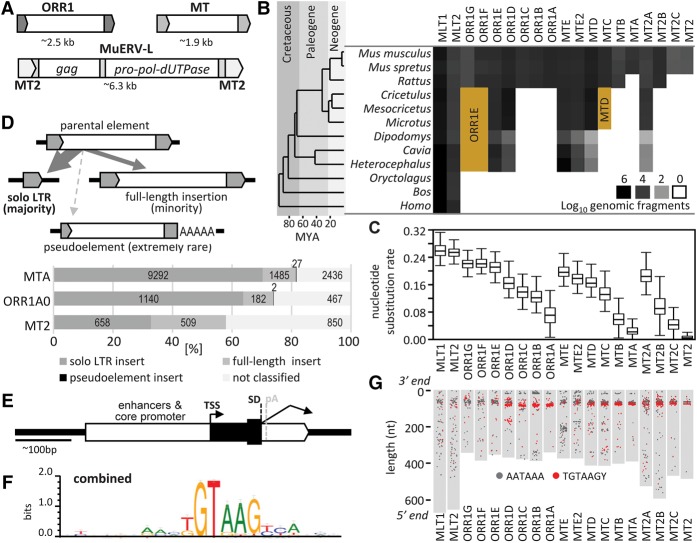
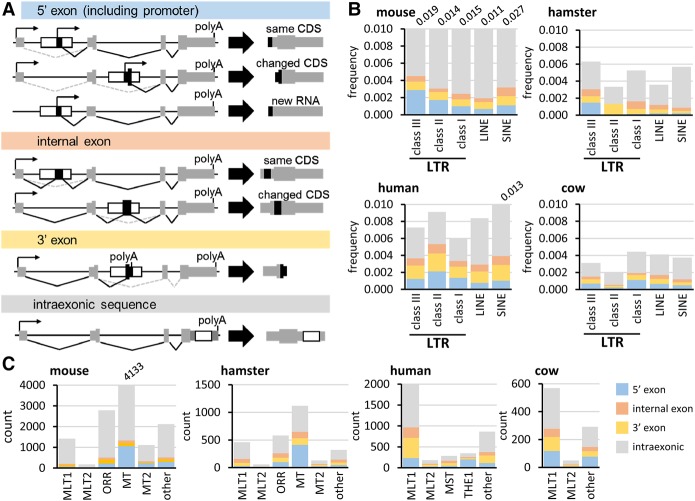
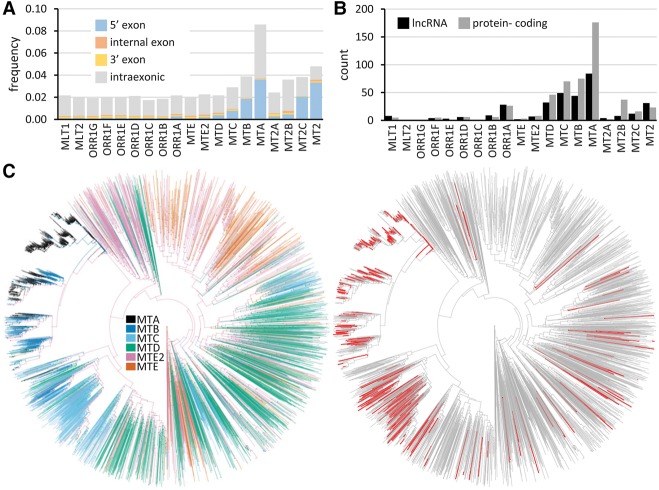
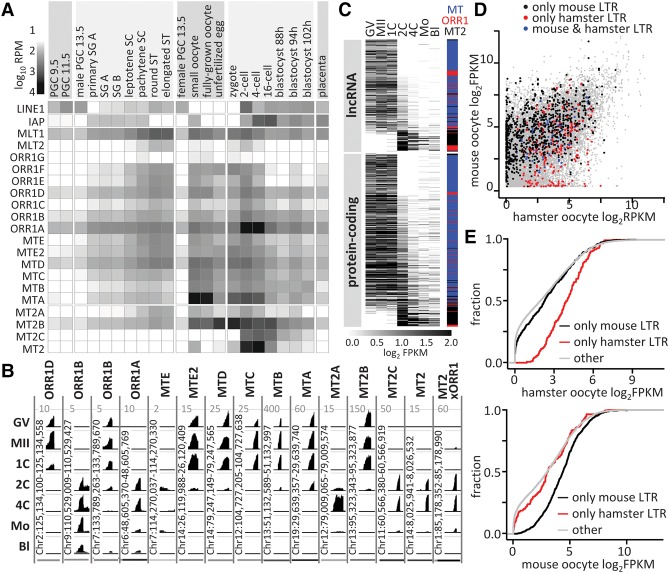
Retrotransposons are "copy-and-paste" insertional mutagens that substantially contribute to mammalian genome content. Retrotransposons often carry long terminal repeats (LTRs) for retrovirus-like reverse transcription and integration into the genome. We report an extraordinary impact of a group of LTRs from the mammalian endogenous retrovirus-related ERVL retrotransposon class on gene expression in the germline and beyond. In mouse, we identified more than 800 LTRs from ORR1, MT, MT2, and MLT families, which resemble mobile gene-remodeling platforms that supply promoters and first exons. The LTR-mediated gene remodeling also extends to hamster, human, and bovine oocytes. The LTRs function in a stage-specific manner during the oocyte-to-embryo transition by activating transcription, altering protein-coding sequences, producing noncoding RNAs, and even supporting evolution of new protein-coding genes. These functions result, for example, in recycling processed pseudogenes into mRNAs or lncRNAs with regulatory roles. The functional potential of the studied LTRs is even higher, because we show that dormant LTR promoter activity can rescue loss of an essential upstream promoter. We also report a novel protein-coding gene evolution-D6Ertd527e-in which an MT LTR provided a promoter and the 5' exon with a functional start codon while the bulk of the protein-coding sequence evolved through a CAG repeat expansion. Altogether, ERVL LTRs provide molecular mechanisms for stochastically scanning, rewiring, and recycling genetic information on an extraordinary scale. ERVL LTRs thus offer means for a comprehensive survey of the genome's expression potential, tightly intertwining with gene expression and evolution in the germline.
© 2017 Franke et al.; Published by Cold Spring Harbor Laboratory Press.
Figures
Similar articles
-
Long non-coding RNAs transcribed by ERV-9 LTR retrotransposon act in cis to modulate long-range LTR enhancer function.Nucleic Acids Res. 2017 May 5;45(8):4479-4492. doi: 10.1093/nar/gkx055. Nucleic Acids Res. 2017. PMID: 28132025 Free PMC article.
-
Repeated recruitment of LTR retrotransposons as promoters by the anti-apoptotic locus NAIP during mammalian evolution.PLoS Genet. 2007 Jan 12;3(1):e10. doi: 10.1371/journal.pgen.0030010. Epub 2006 Dec 6. PLoS Genet. 2007. PMID: 17222062 Free PMC article.
-
Retrotransposons and their recognition of pol II promoters: a comprehensive survey of the transposable elements from the complete genome sequence of Schizosaccharomyces pombe.Genome Res. 2003 Sep;13(9):1984-97. doi: 10.1101/gr.1191603. Genome Res. 2003. PMID: 12952871 Free PMC article.
-
Endogenous retroviral LTRs as promoters for human genes: a critical assessment.Gene. 2009 Dec 15;448(2):105-14. doi: 10.1016/j.gene.2009.06.020. Epub 2009 Jul 3. Gene. 2009. PMID: 19577618 Review.
-
Epigenetic Control of Human Endogenous Retrovirus Expression: Focus on Regulation of Long-Terminal Repeats (LTRs).Viruses. 2017 May 31;9(6):130. doi: 10.3390/v9060130. Viruses. 2017. PMID: 28561791 Free PMC article. Review.
Cited by
-
Transcription of Endogenous Retroviruses: Broad and Precise Mechanisms of Control.Viruses. 2024 Aug 17;16(8):1312. doi: 10.3390/v16081312. Viruses. 2024. PMID: 39205286 Free PMC article. Review.
-
Zebrafish transposable elements show extensive diversification in age, genomic distribution, and developmental expression.Genome Res. 2022 Jul;32(7):1408-1423. doi: 10.1101/gr.275655.121. Epub 2022 Jan 5. Genome Res. 2022. PMID: 34987056 Free PMC article.
-
Chromatin remodeling in bovine embryos indicates species-specific regulation of genome activation.Nat Commun. 2020 Sep 17;11(1):4654. doi: 10.1038/s41467-020-18508-3. Nat Commun. 2020. PMID: 32943640 Free PMC article.
-
Evolution and subfamilies of HERVL human endogenous retrovirus.Bioinform Adv. 2024 Jul 30;4(1):vbae110. doi: 10.1093/bioadv/vbae110. eCollection 2024. Bioinform Adv. 2024. PMID: 39139705 Free PMC article.
-
HERVs, immunity, and autoimmunity: understanding the connection.PeerJ. 2019 Apr 5;7:e6711. doi: 10.7717/peerj.6711. eCollection 2019. PeerJ. 2019. PMID: 30984482 Free PMC article.
References
-
- Bénit L, De Parseval N, Casella JF, Callebaut I, Cordonnier A, Heidmann T. 1997. Cloning of a new murine endogenous retrovirus, MuERV-L, with strong similarity to the human HERV-L element and with a gag coding sequence closely related to the Fv1 restriction gene. J Virol 71: 5652–5657. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases