

A defined metabolic state in pre B cells governs B-cell development and is counterbalanced by Swiprosin-2/EFhd1
- PMID: 28524857
- PMCID: PMC5520169
- DOI: 10.1038/cdd.2017.52
A defined metabolic state in pre B cells governs B-cell development and is counterbalanced by Swiprosin-2/EFhd1
Abstract
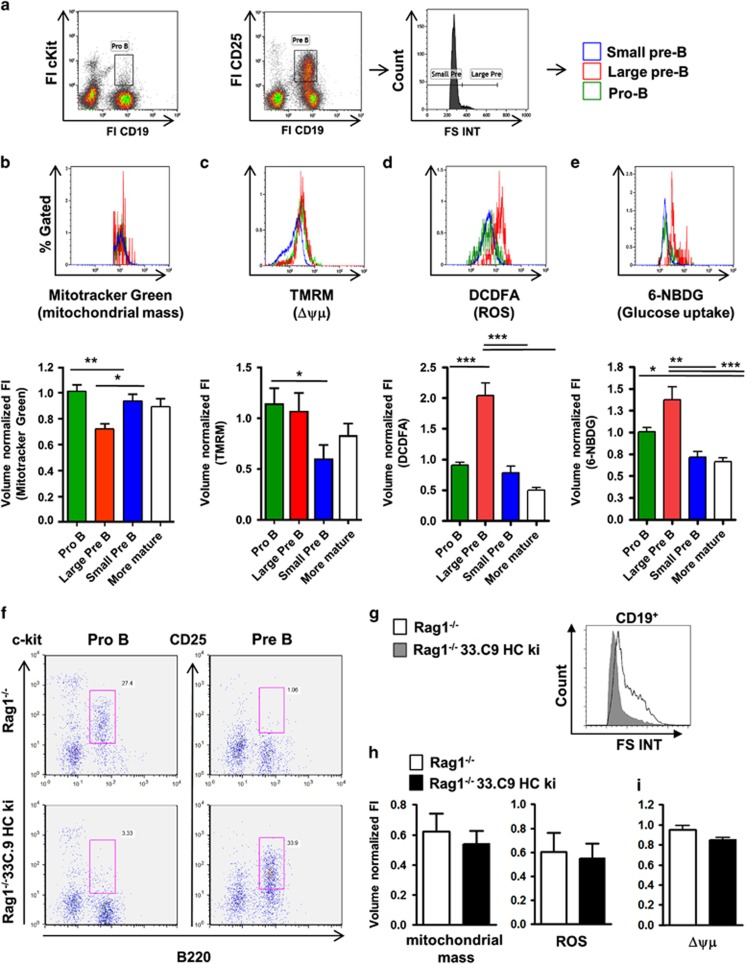
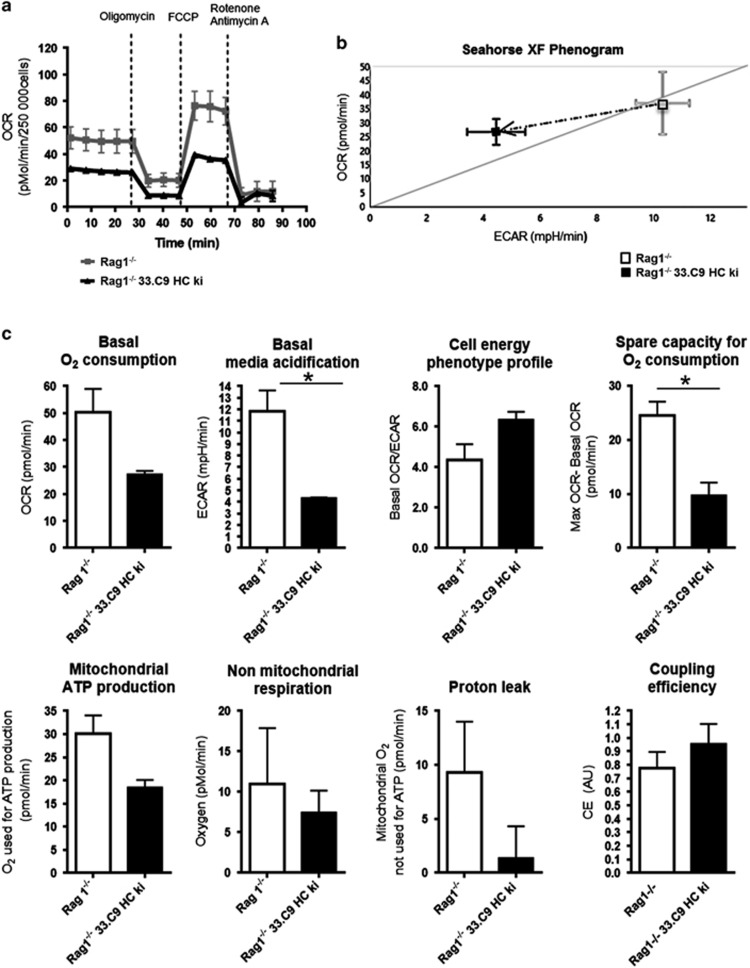
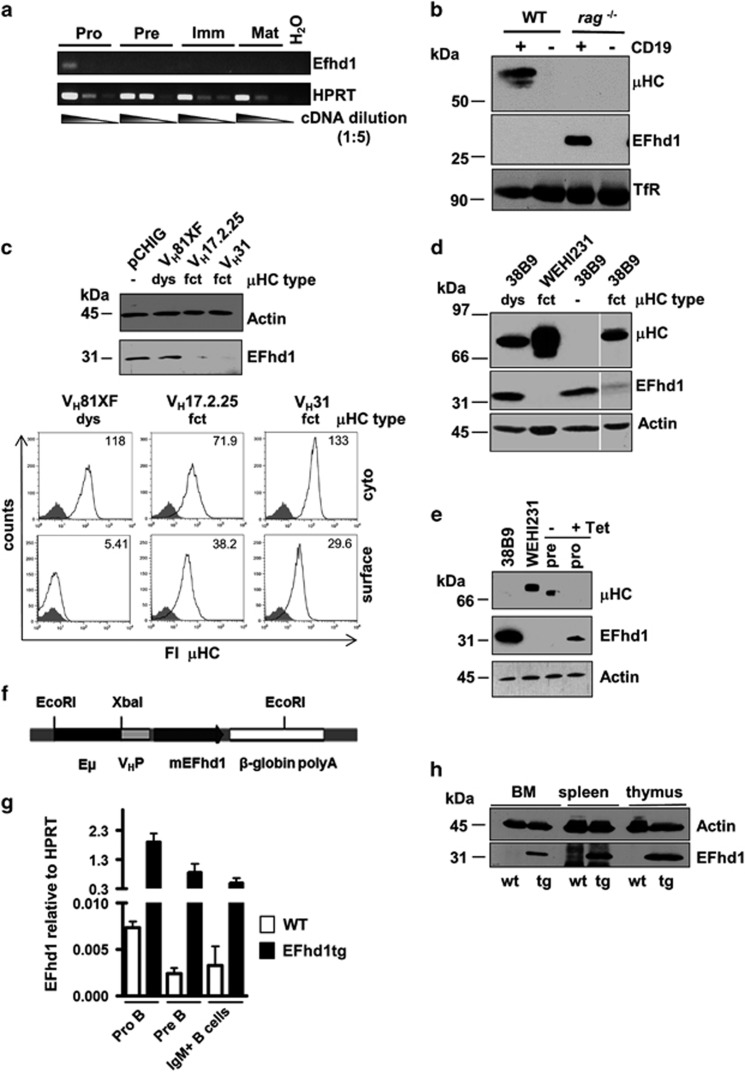
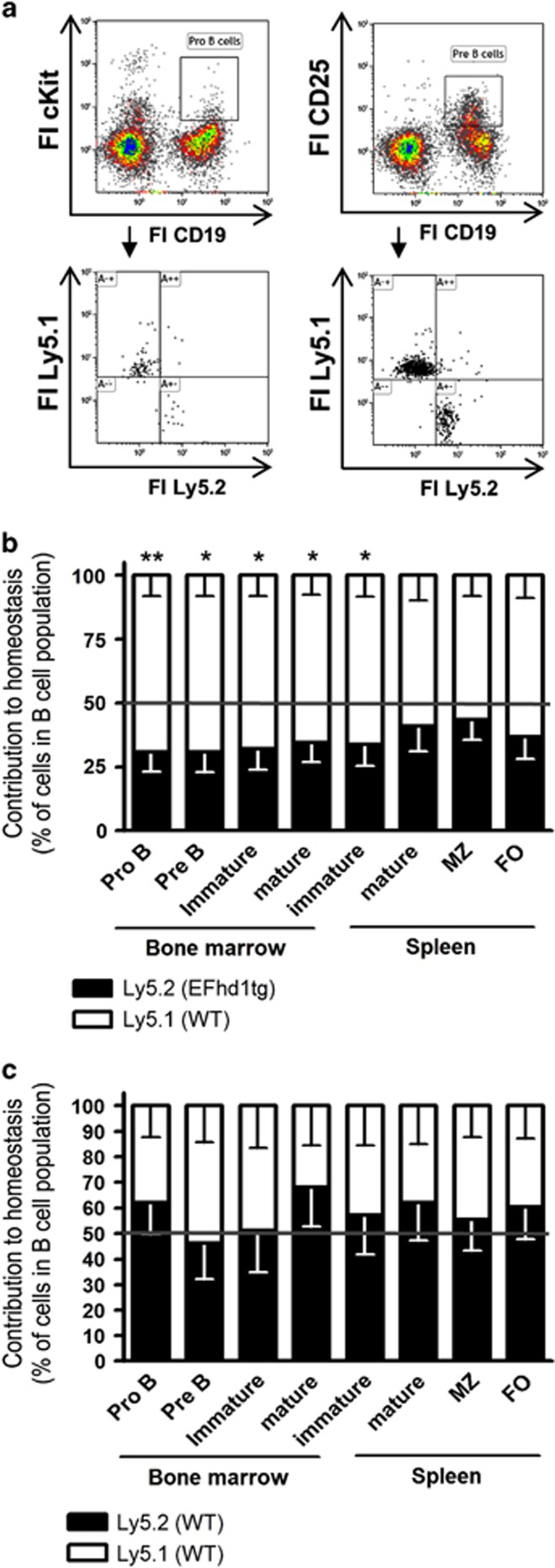
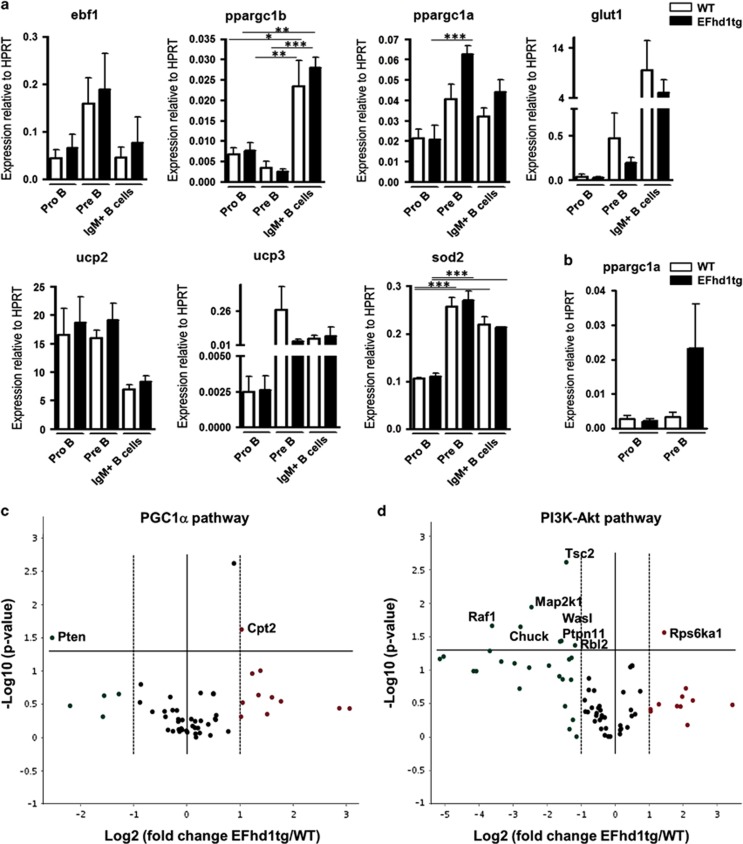
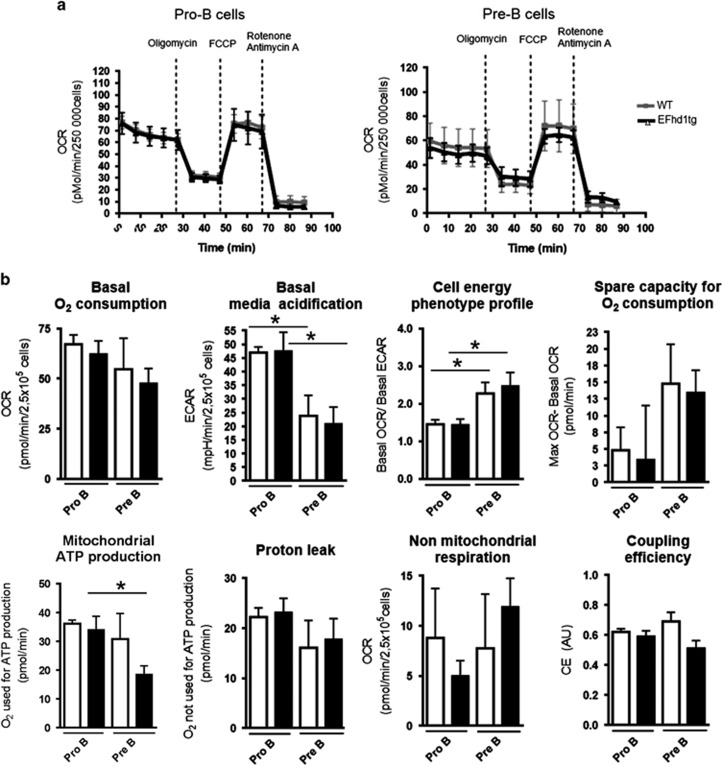
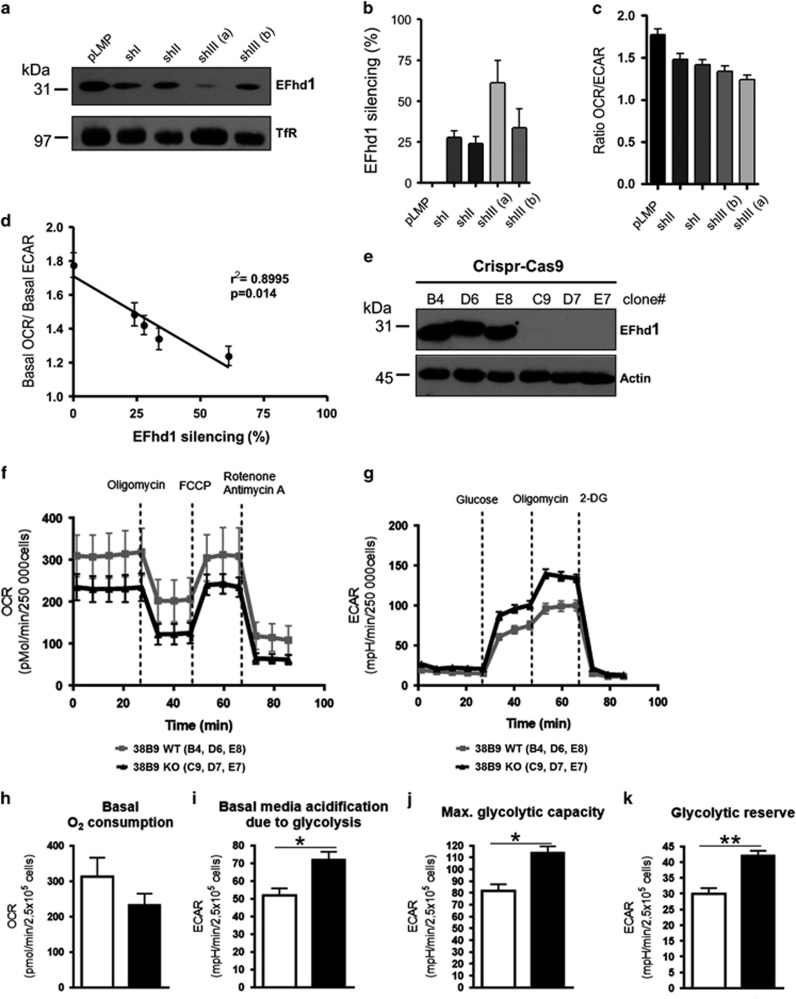
B-cell development in the bone marrow comprises proliferative and resting phases in different niches. We asked whether B-cell metabolism relates to these changes. Compared to pro B and small pre B cells, large pre B cells revealed the highest glucose uptake and ROS but not mitochondrial mass, whereas small pre B cells exhibited the lowest mitochondrial membrane potential. Small pre B cells from Rag1-/-;33.C9 μ heavy chain knock-in mice revealed decreased glycolysis (ECAR) and mitochondrial spare capacity compared to pro B cells from Rag1-/- mice. We were interested in the step regulating this metabolic switch from pro to pre B cells and uncovered that Swiprosin-2/EFhd1, a Ca2+-binding protein of the inner mitochondrial membrane involved in Ca2+-induced mitoflashes, is expressed in pro B cells, but downregulated by surface pre B-cell receptor expression. Knockdown and knockout of EFhd1 in 38B9 pro B cells decreased the oxidative phosphorylation/glycolysis (OCR/ECAR) ratio by increasing glycolysis, glycolytic capacity and reserve. Prolonged expression of EFhd1 in EFhd1 transgenic mice beyond the pro B cell stage increased expression of the mitochondrial co-activator PGC-1α in primary pre B cells, but reduced mitochondrial ATP production at the pro to pre B cell transition in IL-7 cultures. Transgenic EFhd1 expression caused a B-cell intrinsic developmental disadvantage for pro and pre B cells. Hence, coordinated expression of EFhd1 in pro B cells and by the pre BCR regulates metabolic changes and pro/pre B-cell development.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Melchers F. The pre-B-cell receptor: selector of fitting immunoglobulin heavy chains for the B-cell repertoire. Nat Rev Immunol 2005; 5: 578–584. - PubMed
-
- Vettermann C, Jäck HM. The pre-B cell receptor: turning autoreactivity into self-defense. Trends Immunol 2010; 31: 176–183. - PubMed
-
- Young F, Ardman B, Shinkai Y, Lansford R, Blackwell TK, Mendelsohn M et al. Influence of immunoglobulin heavy- and light-chain expression on B-cell differentiation. Genes Dev 1994; 8: 1043–1057. - PubMed
-
- Mombaerts P, Johnson RS, Herrup K, Tonegawa S, Papaioannouo VE. RAG-1-deficient mice have no mature B and T lymphocytes. Cell 1992; 68: 869–877. - PubMed
-
- Guo B, Kato RM, Garcia-Lloret M, Wahl MI, Rawlings DJ. Engagement of the human pre-B cell receptor generates a lipid raft-dependent calcium signaling complex. Immunity 2000; 13: 243–253. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
