

Ubiquitin Modification by the E3 Ligase/ADP-Ribosyltransferase Dtx3L/Parp9
- PMID: 28525742
- PMCID: PMC5556935
- DOI: 10.1016/j.molcel.2017.04.028
Ubiquitin Modification by the E3 Ligase/ADP-Ribosyltransferase Dtx3L/Parp9
Abstract
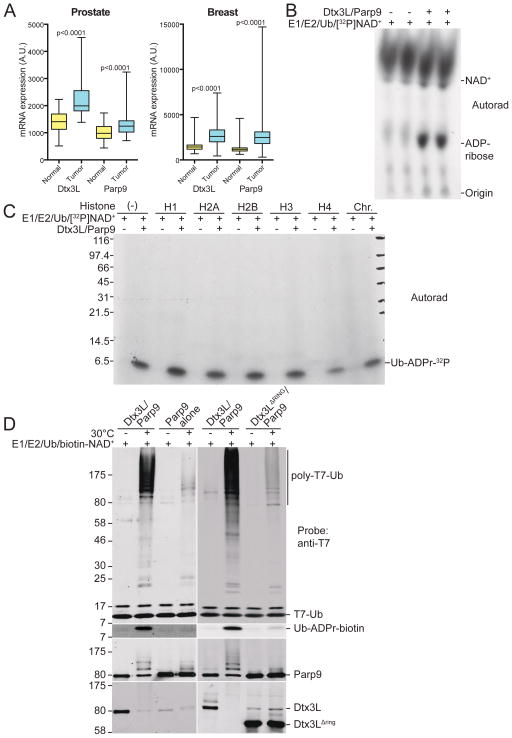
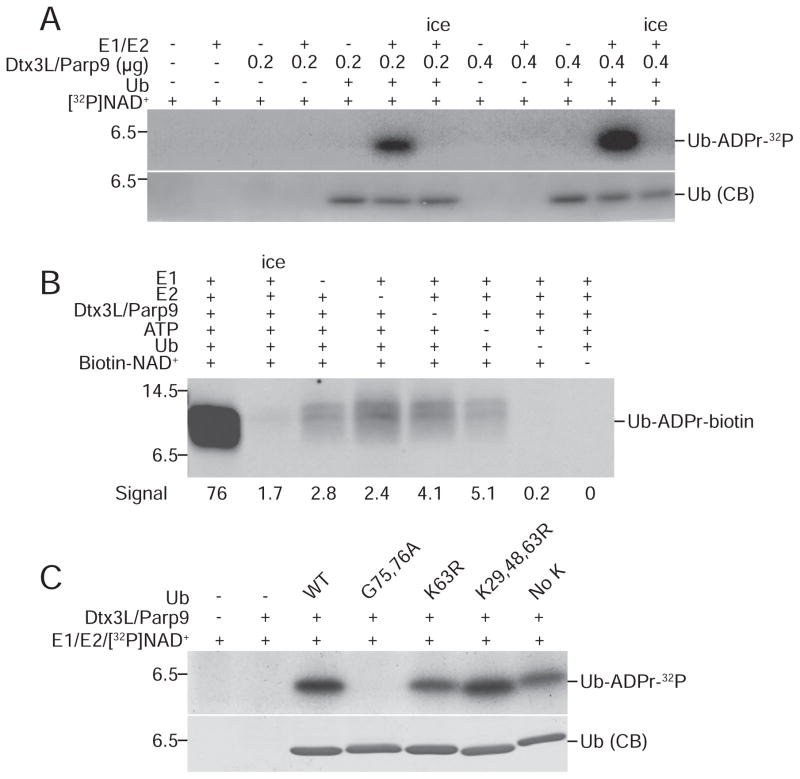
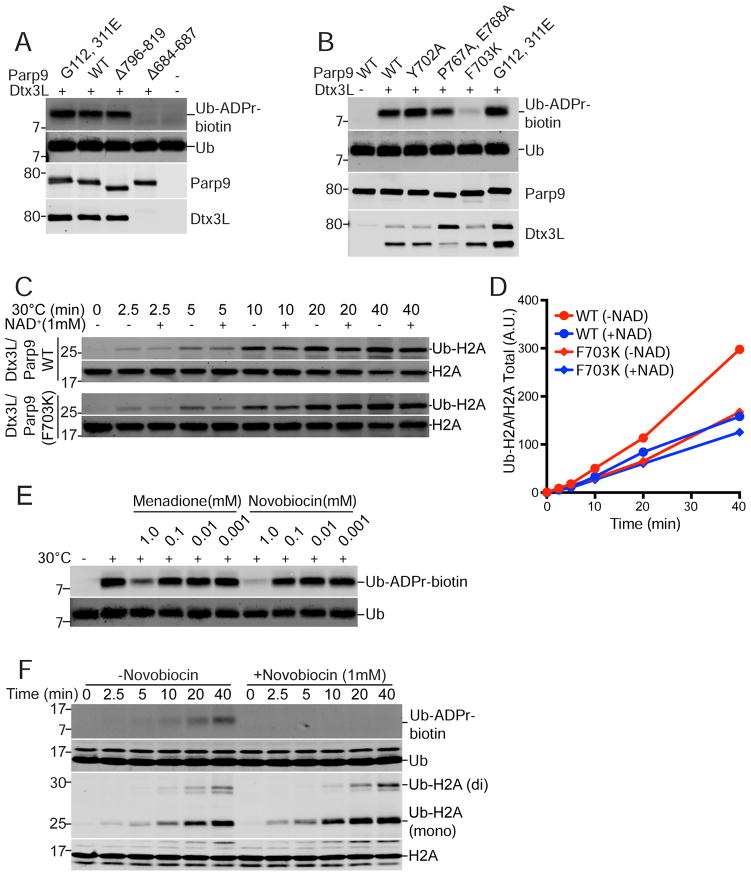
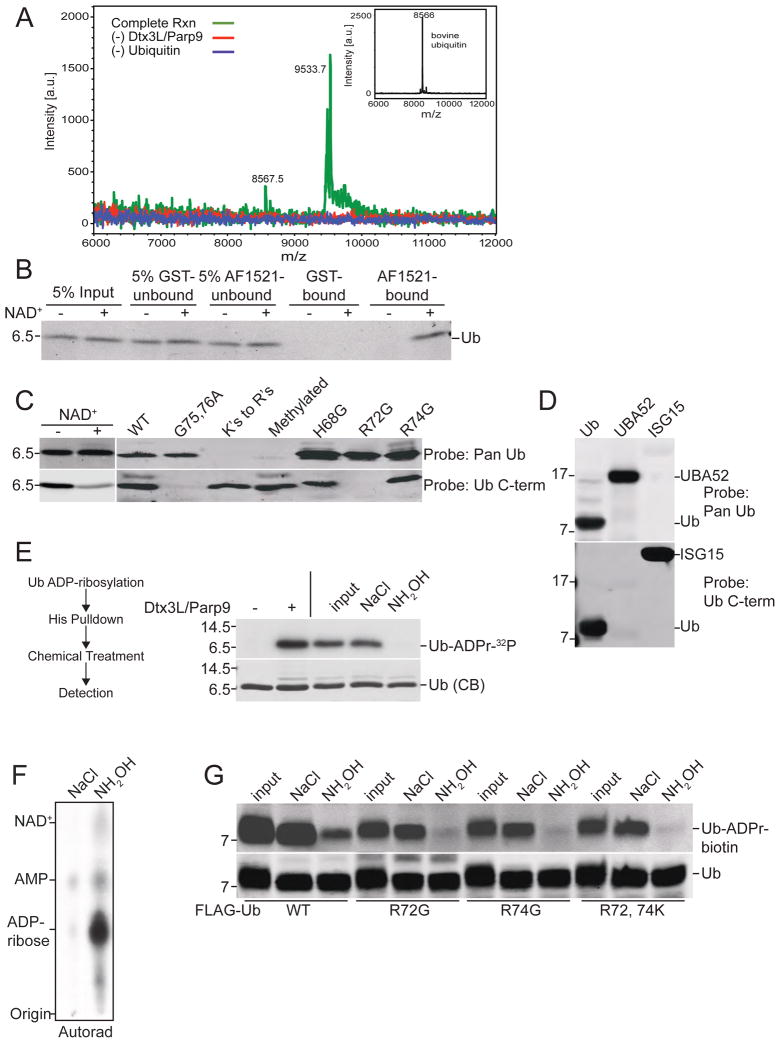
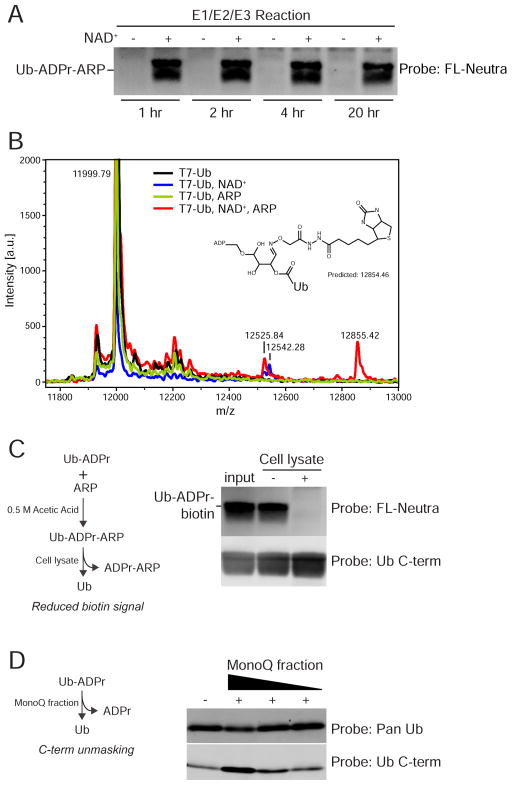
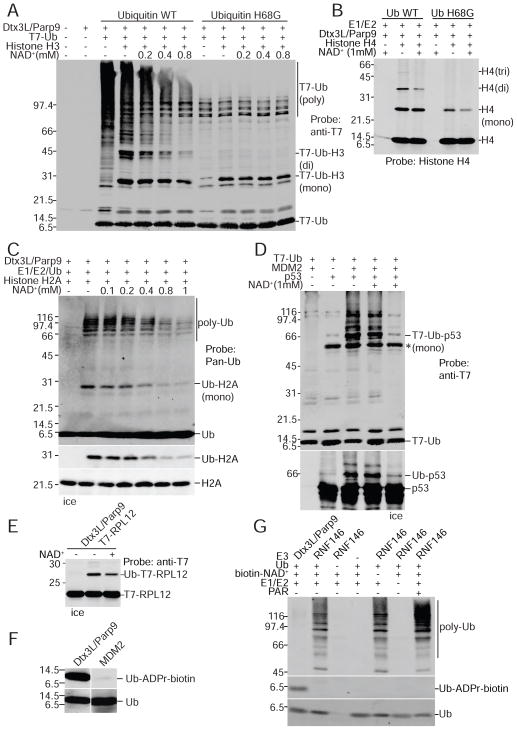
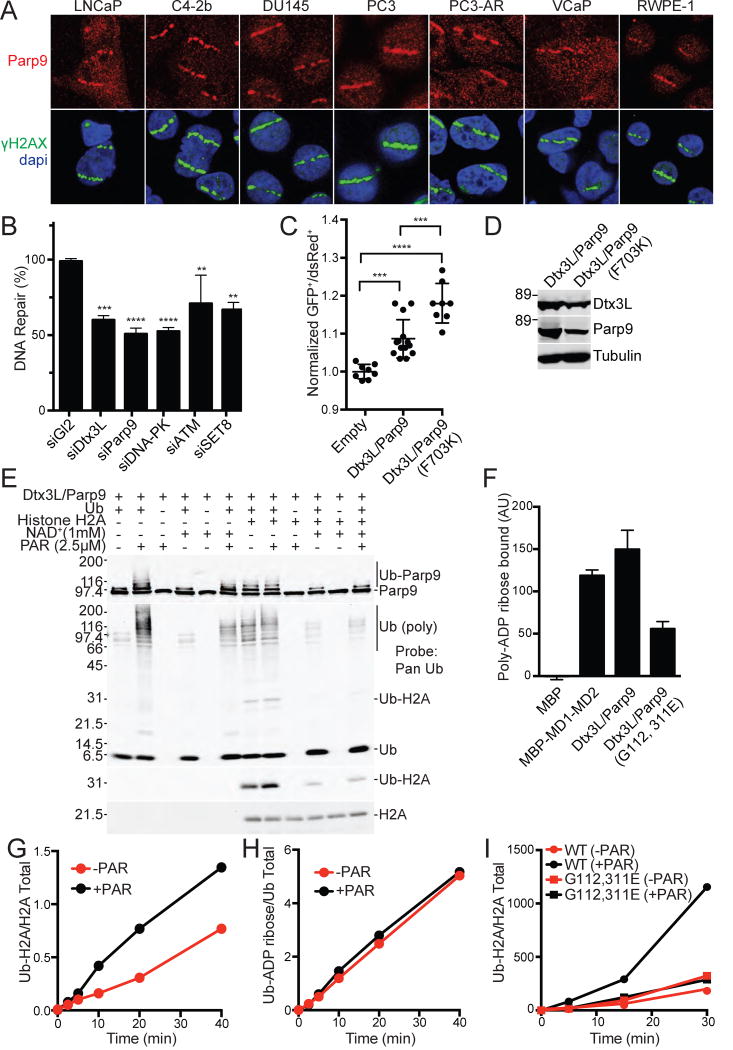
ADP-ribosylation of proteins is emerging as an important regulatory mechanism. Depending on the family member, ADP-ribosyltransferases either conjugate a single ADP-ribose to a target or generate ADP-ribose chains. Here we characterize Parp9, a mono-ADP-ribosyltransferase reported to be enzymatically inactive. Parp9 undergoes heterodimerization with Dtx3L, a histone E3 ligase involved in DNA damage repair. We show that the Dtx3L/Parp9 heterodimer mediates NAD+-dependent mono-ADP-ribosylation of ubiquitin, exclusively in the context of ubiquitin processing by E1 and E2 enzymes. Dtx3L/Parp9 ADP-ribosylates the carboxyl group of Ub Gly76. Because Gly76 is normally used for Ub conjugation to substrates, ADP-ribosylation of the Ub carboxyl terminus precludes ubiquitylation. Parp9 ADP-ribosylation activity therefore restrains the E3 function of Dtx3L. Mutation of the NAD+ binding site in Parp9 increases the DNA repair activity of the heterodimer. Moreover, poly(ADP-ribose) binding to the Parp9 macrodomains increases E3 activity. Dtx3L heterodimerization with Parp9 enables NAD+ and poly(ADP-ribose) regulation of E3 activity.
Copyright © 2017 Elsevier Inc. All rights reserved.
Figures
References
-
- Aguiar RC, Takeyama K, He C, Kreinbrink K, Shipp MA. B-aggressive lymphoma family proteins have unique domains that modulate transcription and exhibit poly(ADP-ribose) polymerase activity. J Biol Chem. 2005;280:33756–33765. - PubMed
-
- Aguiar RC, Yakushijin Y, Kharbanda S, Salgia R, Fletcher JA, Shipp MA. BAL is a novel risk-related gene in diffuse large B-cell lymphomas that enhances cellular migration. Blood. 2000;96:4328–4334. - PubMed
-
- Banasik M, Komura H, Shimoyama M, Ueda K. Specific inhibitors of poly(ADP-ribose) synthetase and mono(ADP-ribosyl)transferase. J Biol Chem. 1992;267:1569–1575. - PubMed
-
- Barkauskaite E, Jankevicius G, Ahel I. Structures and Mechanisms of Enzymes Employed in the Synthesis and Degradation of PARP-Dependent Protein ADP-Ribosylation. Mol Cell. 2015;58:935–946. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
