

Adoptive Transfer of Phosphoantigen-Specific γδ T Cell Subset Attenuates Mycobacterium tuberculosis Infection in Nonhuman Primates
- PMID: 28526681
- PMCID: PMC5557270
- DOI: 10.4049/jimmunol.1602019
Adoptive Transfer of Phosphoantigen-Specific γδ T Cell Subset Attenuates Mycobacterium tuberculosis Infection in Nonhuman Primates
Abstract
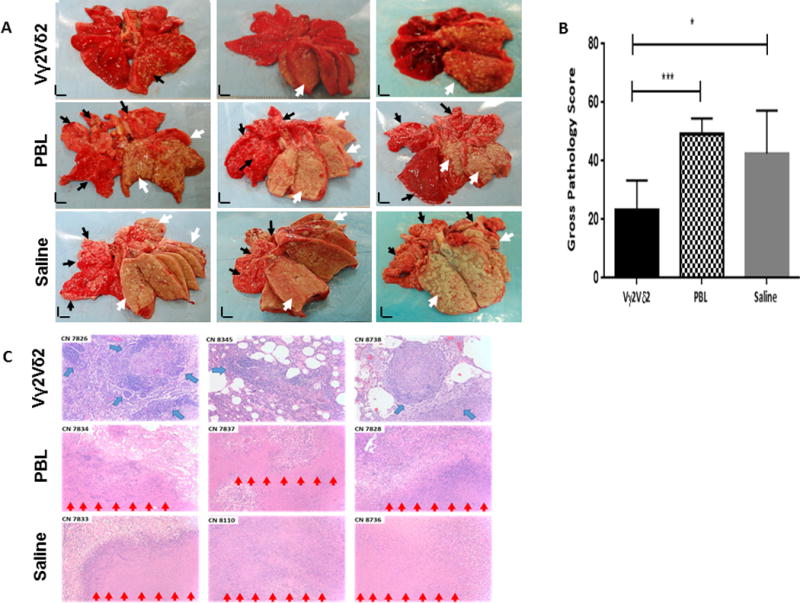
The dominant Vγ2Vδ2 T cell subset recognizes phosphoantigen and exists only in humans and nonhuman primates. Despite the discovery of γδ T cells >30 y ago, a proof-of-concept study has not been done to prove the principle that the Vγ2Vδ2 T cell subset is protective against Mycobacterium tuberculosis and other infections. In this study, we used an adoptive cell-transfer strategy to define the protective role of Vγ2Vδ2 T cells in a primate tuberculosis (TB) model. Vγ2Vδ2 T cells for adoptive transfer displayed central/effector memory and mounted effector functions, including the production of anti-M. tuberculosis cytokines and inhibition of intracellular mycobacteria. They also expressed CXCR3/CCR5/LFA-1 trafficking/tissue-resident phenotypes and consistently trafficked to the airway, where they remained detectable from 6 h through 7 d after adoptive transfer. Interestingly, the test group of macaques receiving transfer of Vγ2Vδ2 T cells at weeks 1 and 3 after high-dose (500 CFU) M. tuberculosis infection exhibited significantly lower levels of M. tuberculosis infection burdens in lung lobes and extrapulmonary organs than did the control groups receiving PBLs or saline. Consistently, adoptive transfer of Vγ2Vδ2 T cells attenuated TB pathology and contained lesions primarily in the infection site of the right caudal lung lobe, with no or reduced TB dissemination to other lobes, spleen, or liver/kidney; in contrast, the controls showed widespread TB dissemination. The proof-of-concept finding supports the view that the dominant Vγ2Vδ2 T cell subset may be included in the rational design of a TB vaccine or host-directed therapy.
Copyright © 2017 by The American Association of Immunologists, Inc.
Figures





References
-
- Wells CD, Cegielski JP, Nelson LJ, Laserson KF, Holtz TH, Finlay A, Castro KG, Weyer K. HIV Infection and Multidrug-Resistant Tuberculosis—The Perfect Storm. Journal of Infectious Diseases. 2007;196:S86–S107. - PubMed
-
- Lawn SD, Zumla AI. Tuberculosis. The Lancet. 2011;378:57–72. - PubMed
-
- Colditz GA, Brewer TF, Berkey CS, Mosteller F. Efficacy of bcg vaccine in the prevention of tuberculosis: Meta-analysis of the published literature. JAMA. 1994;271:698–702. - PubMed
-
- Kaufmann HSE, Gengenbacher M. Recombinant live vaccine candidates against tuberculosis. Current Opinion in Biotechnology. 2012;23:900–907. - PubMed
-
- RODRIGUES LC, DIWAN VK, WHEELER JG. Protective Effect of BCG against Tuberculous Meningitis and Miliary Tuberculosis: A Meta-Analysis. International Journal of Epidemiology. 1993;22:1154–1158. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
