

Mining for Micropeptides
- PMID: 28528987
- PMCID: PMC5565689
- DOI: 10.1016/j.tcb.2017.04.006
Mining for Micropeptides
Abstract
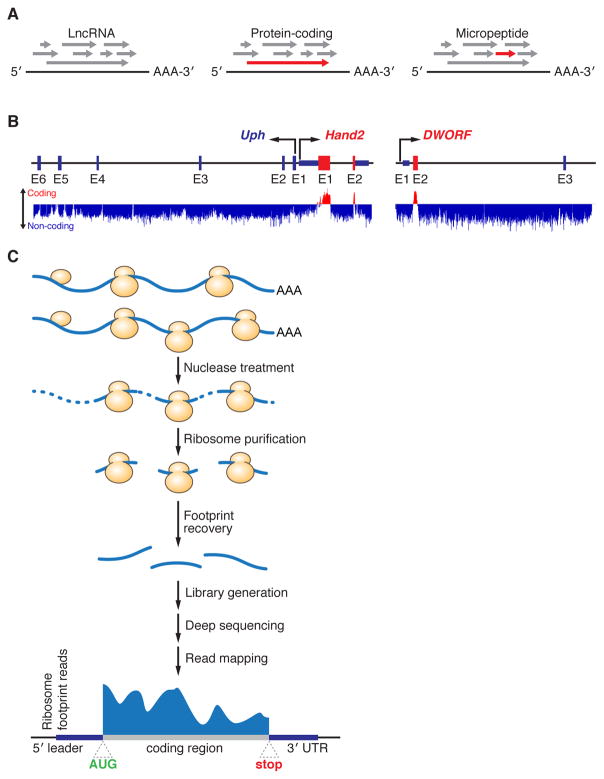
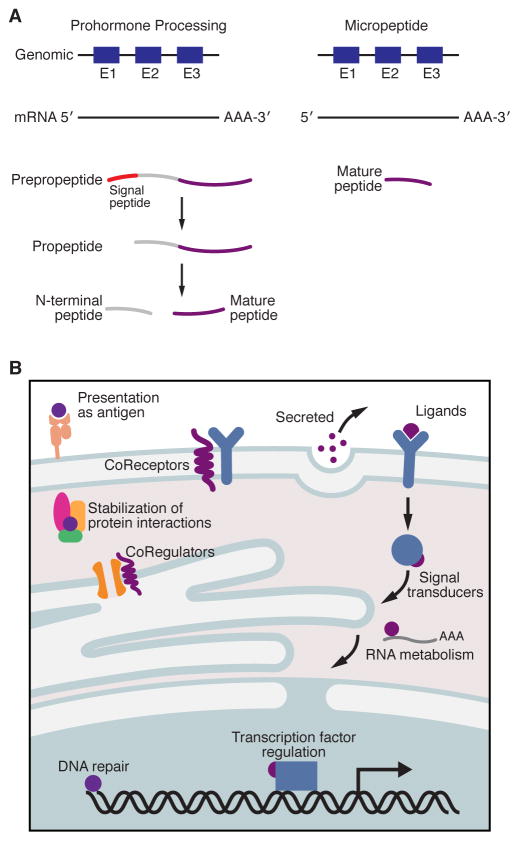
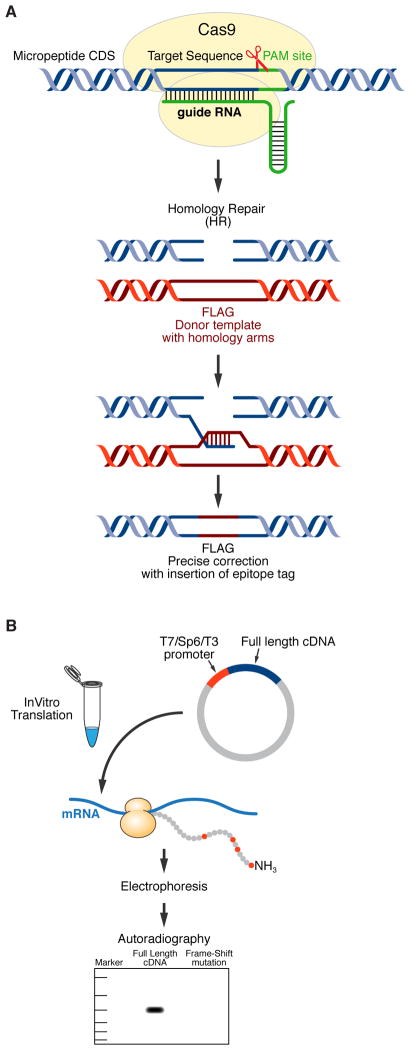
Advances in computational biology and large-scale transcriptome analyses have revealed that a much larger portion of the genome is transcribed than was previously recognized, resulting in the production of a diverse population of RNA molecules with both protein-coding and noncoding potential. Emerging evidence indicates that several RNA molecules have been mis-annotated as noncoding and in fact harbor short open reading frames (sORFs) that encode functional peptides and that have evaded detection until now due to their small size. sORF-encoded peptides (SEPs), or micropeptides, have been shown to have important roles in fundamental biological processes and in the maintenance of cellular homeostasis. These small proteins can act independently, for example as ligands or signaling molecules, or they can exert their biological functions by engaging with and modulating larger regulatory proteins. Given their small size, micropeptides may be uniquely suited to fine-tune complex biological systems.
Keywords: bioactive peptide; micropeptide; ncRNA; short open reading frame.
Copyright © 2017 Elsevier Ltd. All rights reserved.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
