

pSILAC mass spectrometry reveals ZFP91 as IMiD-dependent substrate of the CRL4CRBN ubiquitin ligase
- PMID: 28530236
- PMCID: PMC5458144
- DOI: 10.1038/ncomms15398
pSILAC mass spectrometry reveals ZFP91 as IMiD-dependent substrate of the CRL4CRBN ubiquitin ligase
Abstract
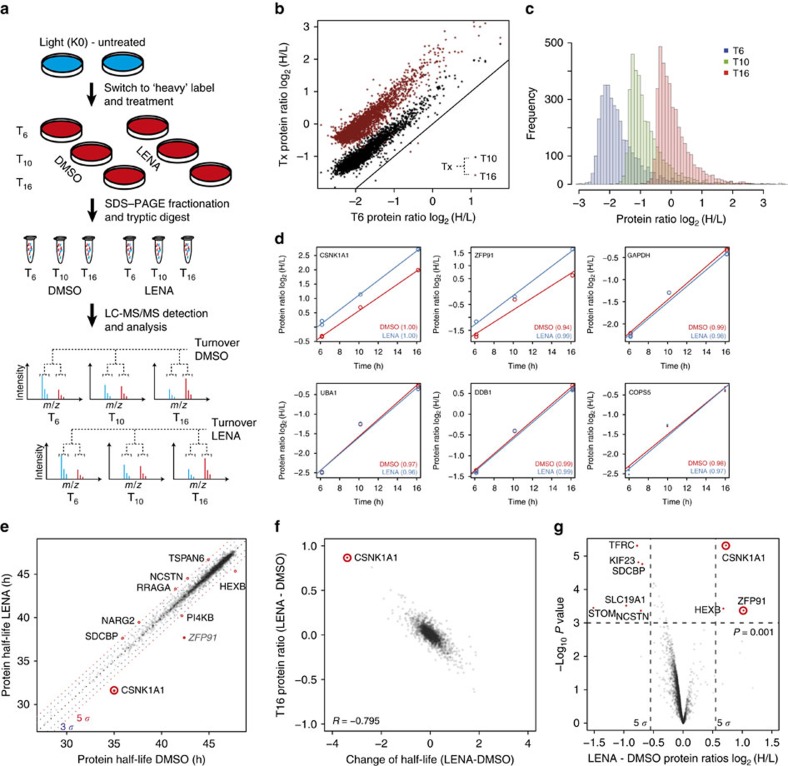
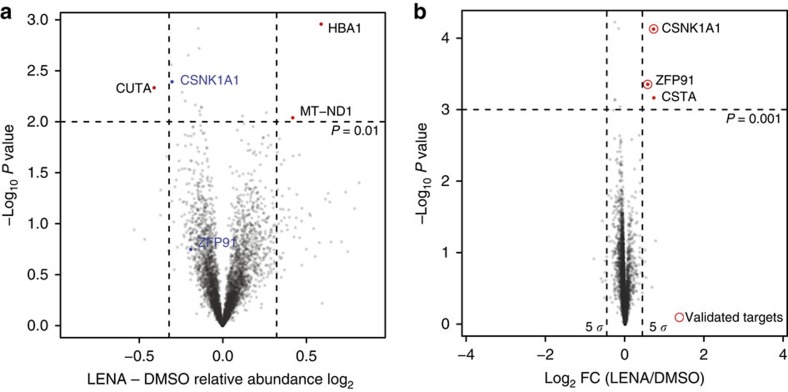
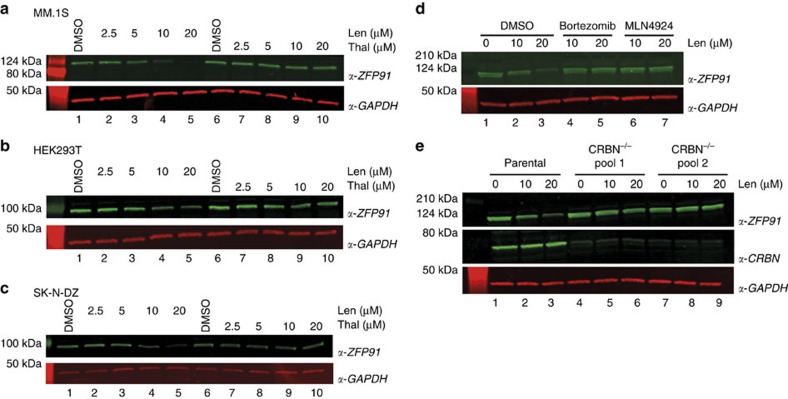
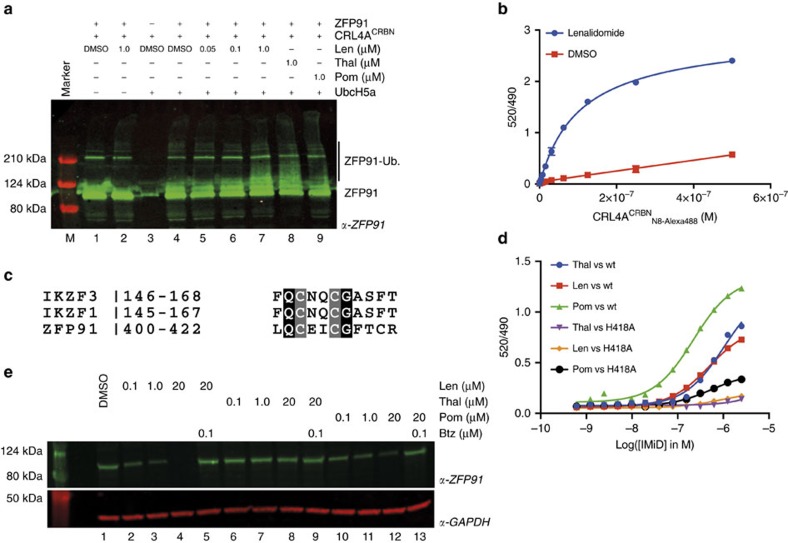
Thalidomide and its derivatives lenalidomide and pomalidomide (IMiDs) are effective treatments of haematologic malignancies. It was shown that IMiDs impart gain-of-function properties to the CUL4-RBX1-DDB1-CRBN (CRL4CRBN) ubiquitin ligase that enable binding, ubiquitination and degradation of key therapeutic targets such as IKZF1, IKZF3 and CSNK1A1. While these substrates have been implicated as efficacy targets in multiple myeloma (MM) and 5q deletion associated myelodysplastic syndrome (del(5q)-MDS), other targets likely exist. Using a pulse-chase SILAC mass spectrometry-based proteomics approach, we demonstrate that lenalidomide induces the ubiquitination and degradation of ZFP91. We establish ZFP91 as a bona fide IMiD-dependent CRL4CRBN substrate and further show that ZFP91 harbours a zinc finger (ZnF) motif, related to the IKZF1/3 ZnF, critical for IMiD-dependent CRBN binding. These findings demonstrate that single time point pulse-chase SILAC mass spectrometry-based proteomics (pSILAC MS) is a sensitive approach for target identification of small molecules inducing selective protein degradation.
Conflict of interest statement
The authors declare no competing financial interests.
Figures

References
-
- Komander D. & Rape M. The ubiquitin code. Annu. Rev. Biochem. 81, 203–229 (2012). - PubMed
-
- O'Connell B. C. & Harper J. W. Ubiquitin proteasome system (UPS): what can chromatin do for you? Curr. Opin. Cell Biol. 19, 206–214 (2007). - PubMed
-
- Hershko A. & Ciechanover A. The ubiquitin system. Annu. Rev. Biochem. 67, 425–479 (1998). - PubMed
-
- Deshaies R. J. & Joazeiro C. A. P. RING domain E3 ubiquitin ligases. Annu. Rev. Biochem. 78, 399–434 (2009). - PubMed
-
- Singhal S. et al. Antitumor activity of thalidomide in refractory multiple myeloma. N. Engl. J. Med. 341, 1565–1571 (1999). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
