

Multi-virion infectious units arise from free viral particles in an enveloped virus
- PMID: 28530650
- PMCID: PMC5447809
- DOI: 10.1038/nmicrobiol.2017.78
Multi-virion infectious units arise from free viral particles in an enveloped virus
Abstract
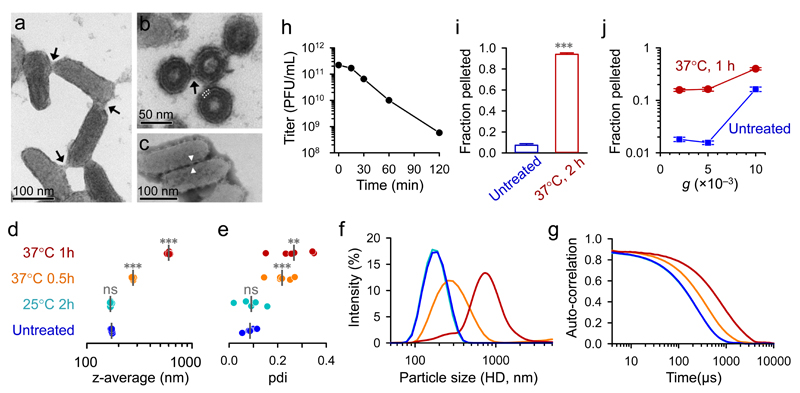
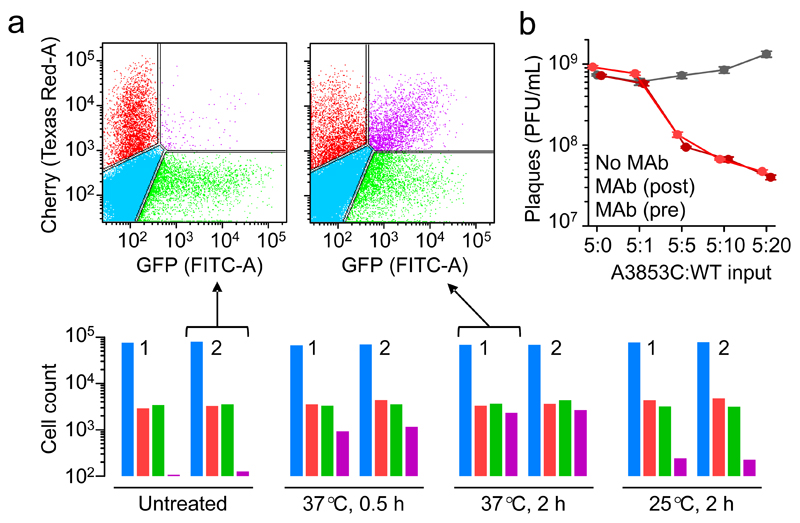
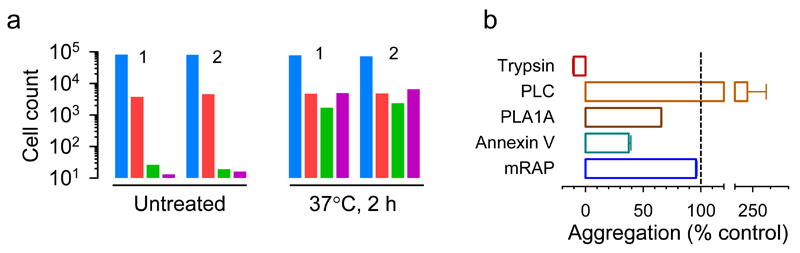
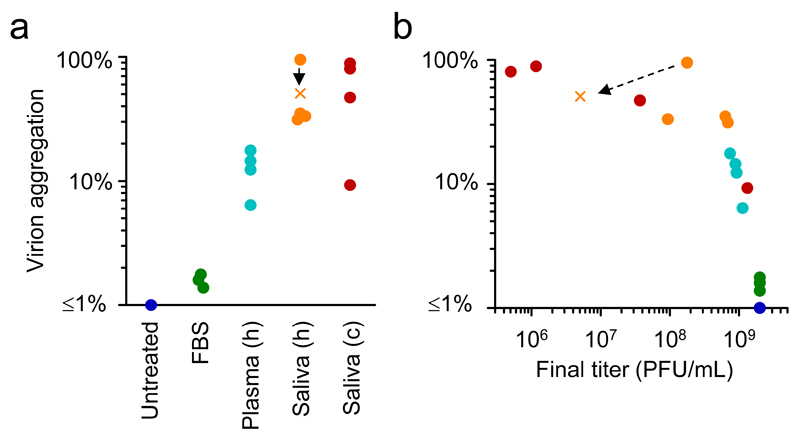
Many animal viruses are enveloped in a lipid bilayer taken up from cellular membranes. Because viral surface proteins bind to these membranes to initiate infection, we hypothesized that free virions may also be capable of interacting with the envelopes of other virions extracellularly. Here, we demonstrate this hypothesis in the vesicular stomatitis virus (VSV), a prototypic negative-strand RNA virus composed of an internal ribonucleocapsid, a matrix protein and an external envelope1. Using microscopy, dynamic light scattering, differential centrifugation and flow cytometry, we show that free viral particles can spontaneously aggregate into multi-virion infectious units. We also show that, following establishment of these contacts, different viral genetic variants are co-transmitted to the same target cell. Furthermore, virion-virion binding can determine key aspects of viral fitness such as antibody escape. In purified virions, this process is driven by protein-lipid interactions probably involving the VSV surface glycoprotein and phosphatidylserine. Whereas we found that multi-virion complexes occurred unfrequently in standard cell cultures, they were abundant in other fluids such as saliva, a natural VSV shedding route2. Our findings contrast with the commonly accepted perception of virions as passive propagules and show the ability of enveloped viruses to establish collective infectious units, which could in turn facilitate the evolution of virus-virus interactions and of social-like traits3.
Conflict of interest statement
Figures
Similar articles
-
Collective Viral Spread Mediated by Virion Aggregates Promotes the Evolution of Defective Interfering Particles.mBio. 2020 Jan 7;11(1):e02156-19. doi: 10.1128/mBio.02156-19. mBio. 2020. PMID: 31911487 Free PMC article.
-
A confocal and electron microscopic comparison of interferon beta-induced changes in vesicular stomatitis virus infection of neuroblastoma and nonneuronal cells.DNA Cell Biol. 2010 Mar;29(3):103-20. doi: 10.1089/dna.2009.0963. DNA Cell Biol. 2010. PMID: 20113203 Free PMC article.
-
A GP64-null baculovirus pseudotyped with vesicular stomatitis virus G protein.J Virol. 2001 Mar;75(6):2544-56. doi: 10.1128/JVI.75.6.2544-2556.2001. J Virol. 2001. PMID: 11222677 Free PMC article.
-
Molecular Mechanism of Externalization of Phosphatidylserine on the Surface of Ebola Virus Particles.DNA Cell Biol. 2019 Feb;38(2):115-120. doi: 10.1089/dna.2018.4485. Epub 2019 Jan 7. DNA Cell Biol. 2019. PMID: 30615471 Review.
-
Virus maturation by budding.Microbiol Mol Biol Rev. 1998 Dec;62(4):1171-90. doi: 10.1128/MMBR.62.4.1171-1190.1998. Microbiol Mol Biol Rev. 1998. PMID: 9841669 Free PMC article. Review.
Cited by
-
Collective Viral Spread Mediated by Virion Aggregates Promotes the Evolution of Defective Interfering Particles.mBio. 2020 Jan 7;11(1):e02156-19. doi: 10.1128/mBio.02156-19. mBio. 2020. PMID: 31911487 Free PMC article.
-
The evolution of collective infectious units in viruses.Virus Res. 2019 May;265:94-101. doi: 10.1016/j.virusres.2019.03.013. Epub 2019 Mar 17. Virus Res. 2019. PMID: 30894320 Free PMC article.
-
Why are viral genomes so fragile? The bottleneck hypothesis.PLoS Comput Biol. 2021 Jul 8;17(7):e1009128. doi: 10.1371/journal.pcbi.1009128. eCollection 2021 Jul. PLoS Comput Biol. 2021. PMID: 34237053 Free PMC article.
-
Neuraminidase Activity Modulates Cellular Coinfection during Influenza A Virus Multicycle Growth.mBio. 2023 Jun 27;14(3):e0359122. doi: 10.1128/mbio.03591-22. Epub 2023 Apr 20. mBio. 2023. PMID: 37078858 Free PMC article.
-
The kinetics of SARS-CoV-2 infection based on a human challenge study.Proc Natl Acad Sci U S A. 2024 Nov 12;121(46):e2406303121. doi: 10.1073/pnas.2406303121. Epub 2024 Nov 7. Proc Natl Acad Sci U S A. 2024. PMID: 39508770 Free PMC article.
References
-
- Letchworth GJ, Rodríguez LL, Del Cbarrera J. Vesicular stomatitis. Vet J. 1999;157:239–260. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
