

A Japanese Encephalitis Virus Vaccine Inducing Antibodies Strongly Enhancing In Vitro Infection Is Protective in Pigs
- PMID: 28531165
- PMCID: PMC5454436
- DOI: 10.3390/v9050124
A Japanese Encephalitis Virus Vaccine Inducing Antibodies Strongly Enhancing In Vitro Infection Is Protective in Pigs
Abstract
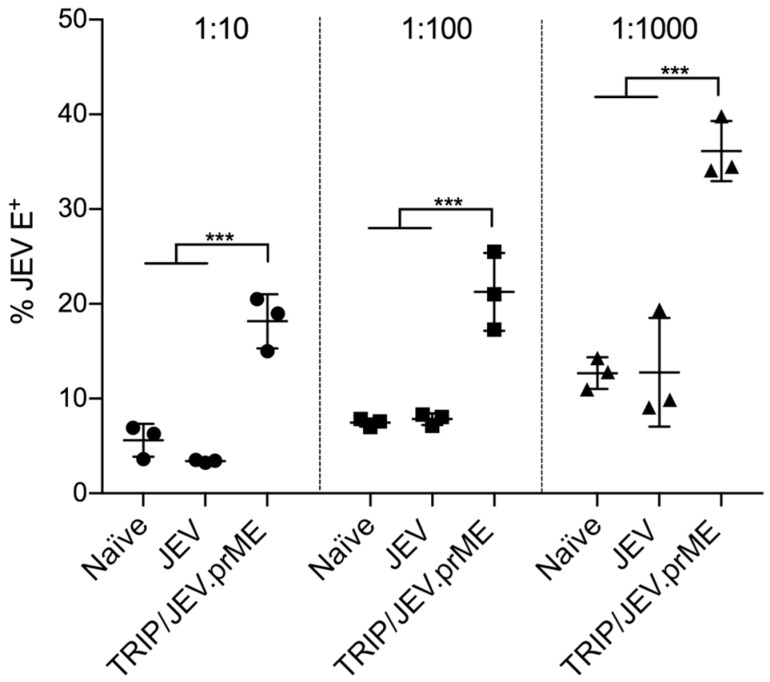
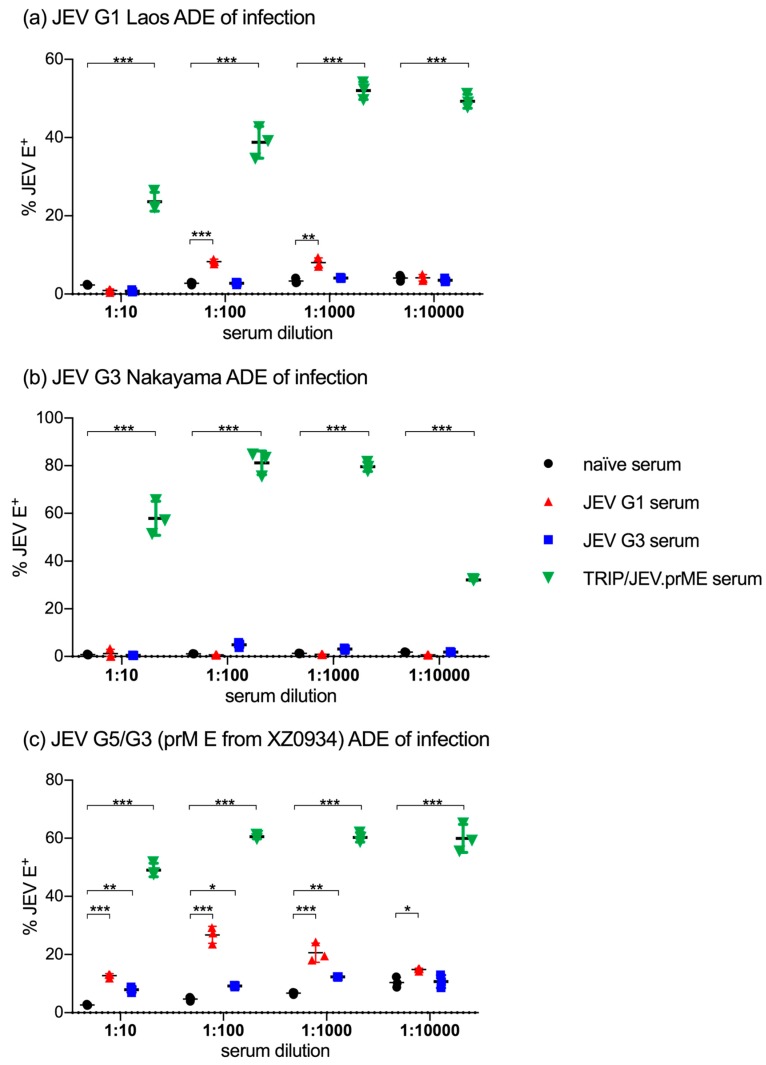
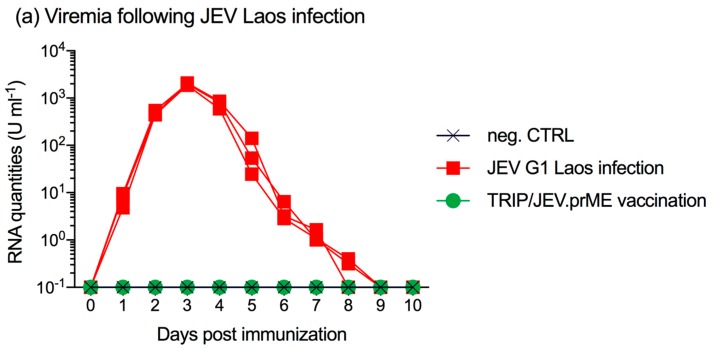
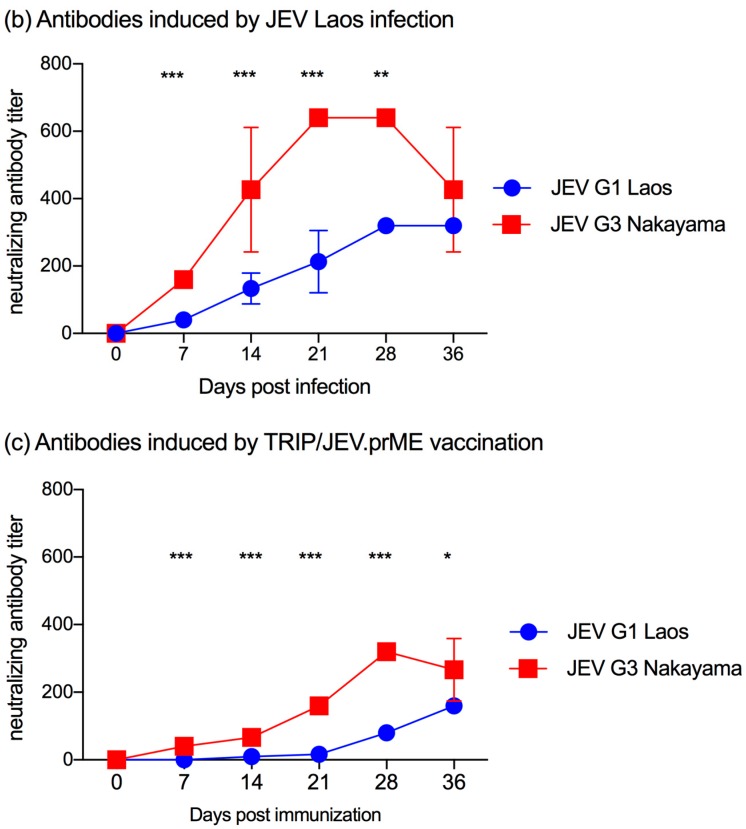
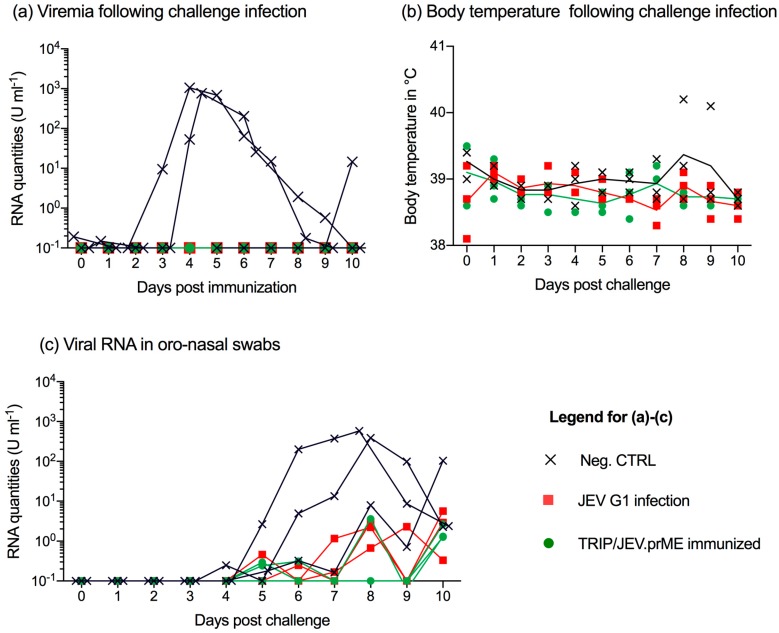
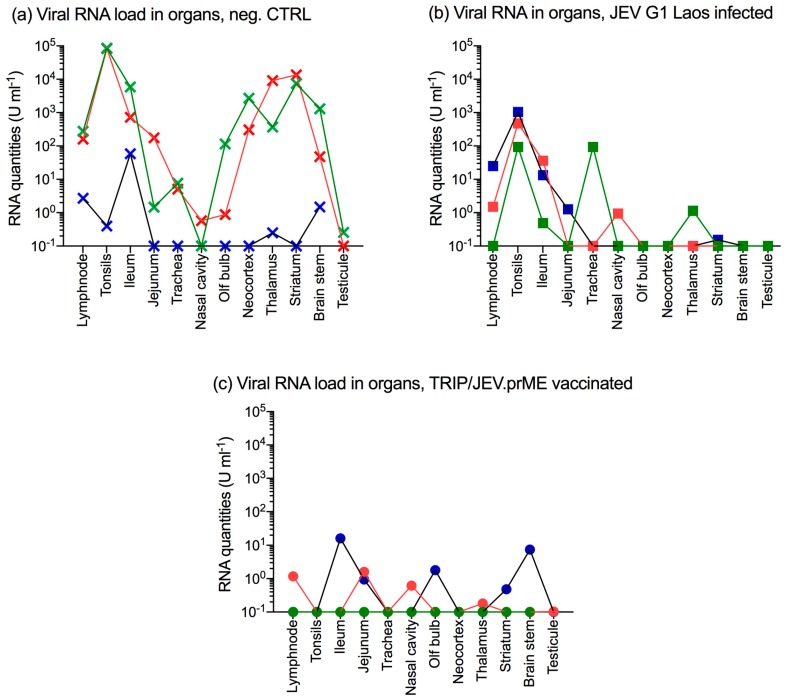
The Japanese encephalitis virus (JEV) is responsible for zoonotic severe viral encephalitis transmitted by Culex mosquitoes. Although birds are reservoirs, pigs play a role as amplifying hosts, and are affected in particular through reproductive failure. Here, we show that a lentiviral JEV vector, expressing JEV prM and E proteins (TRIP/JEV.prME), but not JEV infection induces strong antibody-dependent enhancement (ADE) activities for infection of macrophages. Such antibodies strongly promoted infection via Fc receptors. ADE was found at both neutralizing and non-neutralizing serum dilutions. Nevertheless, in vivo JEV challenge of pigs demonstrated comparable protection induced by the TRIP/JEV.prME vaccine or heterologous JEV infection. Thus, either ADE antibodies cause no harm in the presence of neutralizing antibodies or may even have protective effects in vivo in pigs. Additionally, we found that both pre-infected and vaccinated pigs were not fully protected as low levels of viral RNA were found in lymphoid and nervous system tissue in some animals. Strikingly, the virus from the pre-infection persisted in the tonsils throughout the experiment. Finally, despite the vaccination challenge, viral RNA was detected in the oronasal swabs in all vaccinated pigs. These latter data are relevant when JEV vaccination is employed in pigs.
Keywords: Fc receptor; Japanese encephalitis virus; antibody-dependent enhancement of infection; lentiviral vector vaccine; mucosal virus shedding; persistence; vaccine-induced protection.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Impoinvil D.E., Baylis M., Solomon T. Japanese encephalitis: On the One Health agenda. Curr. Top. Microbiol. Immunol. 2013;365:205–247. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
