

Evolutionary forces affecting synonymous variations in plant genomes
- PMID: 28531201
- PMCID: PMC5460877
- DOI: 10.1371/journal.pgen.1006799
Evolutionary forces affecting synonymous variations in plant genomes
Abstract
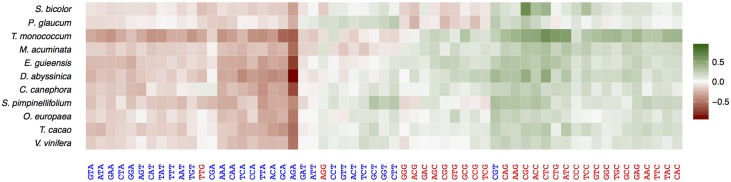
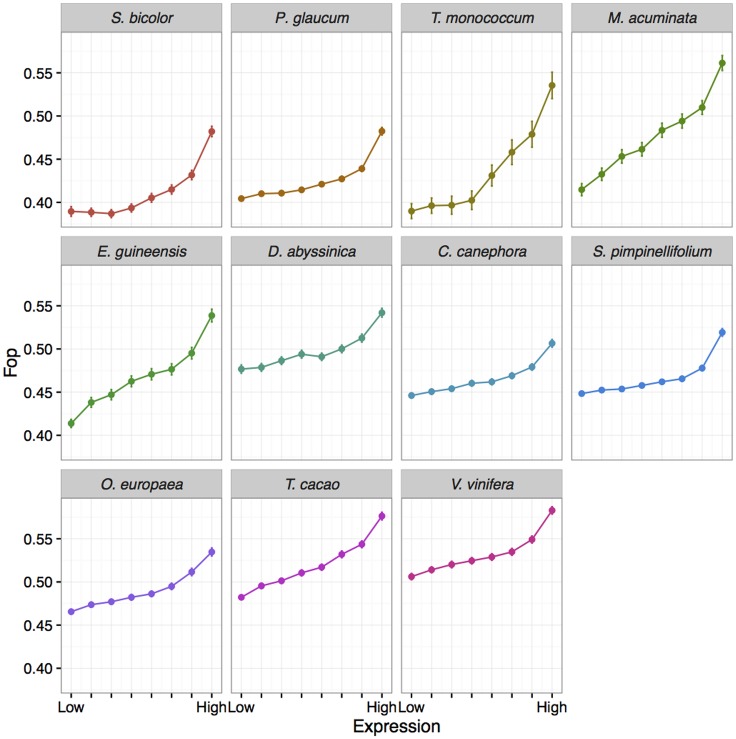
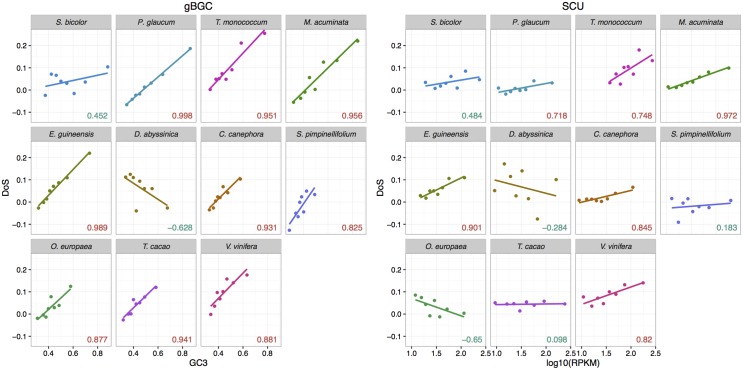
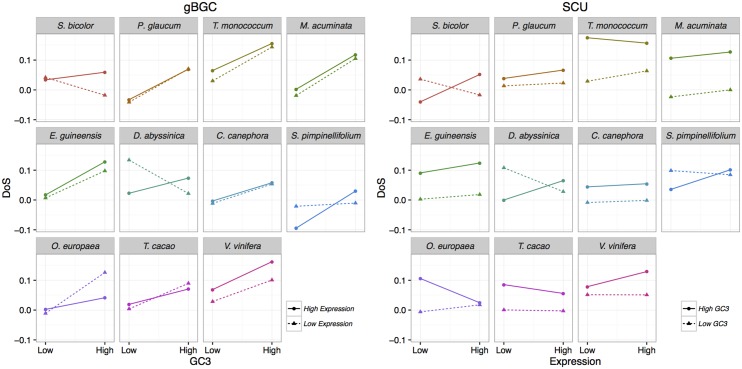
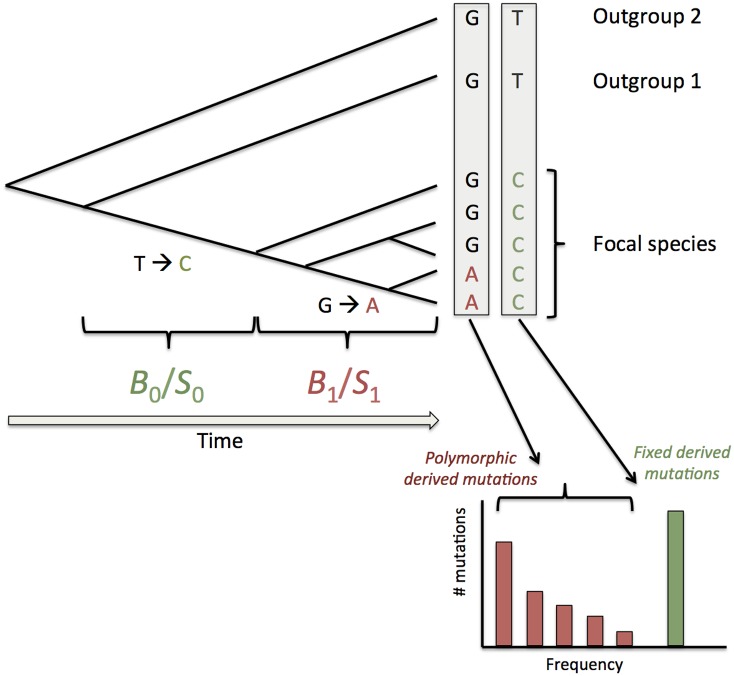
Base composition is highly variable among and within plant genomes, especially at third codon positions, ranging from GC-poor and homogeneous species to GC-rich and highly heterogeneous ones (particularly Monocots). Consequently, synonymous codon usage is biased in most species, even when base composition is relatively homogeneous. The causes of these variations are still under debate, with three main forces being possibly involved: mutational bias, selection and GC-biased gene conversion (gBGC). So far, both selection and gBGC have been detected in some species but how their relative strength varies among and within species remains unclear. Population genetics approaches allow to jointly estimating the intensity of selection, gBGC and mutational bias. We extended a recently developed method and applied it to a large population genomic dataset based on transcriptome sequencing of 11 angiosperm species spread across the phylogeny. We found that at synonymous positions, base composition is far from mutation-drift equilibrium in most genomes and that gBGC is a widespread and stronger process than selection. gBGC could strongly contribute to base composition variation among plant species, implying that it should be taken into account in plant genome analyses, especially for GC-rich ones.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures


References
-
- Wright SI, Yau CB, Looseley M, Meyers BC (2004) Effects of gene expression on molecular evolution in Arabidopsis thaliana and Arabidopsis lyrata. Molecular Biology & Evolution 21: 1719–1726. - PubMed
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
