

Structural basis for substrate binding and catalytic mechanism of a human RNA:m5C methyltransferase NSun6
- PMID: 28531330
- PMCID: PMC5499824
- DOI: 10.1093/nar/gkx473
Structural basis for substrate binding and catalytic mechanism of a human RNA:m5C methyltransferase NSun6
Abstract
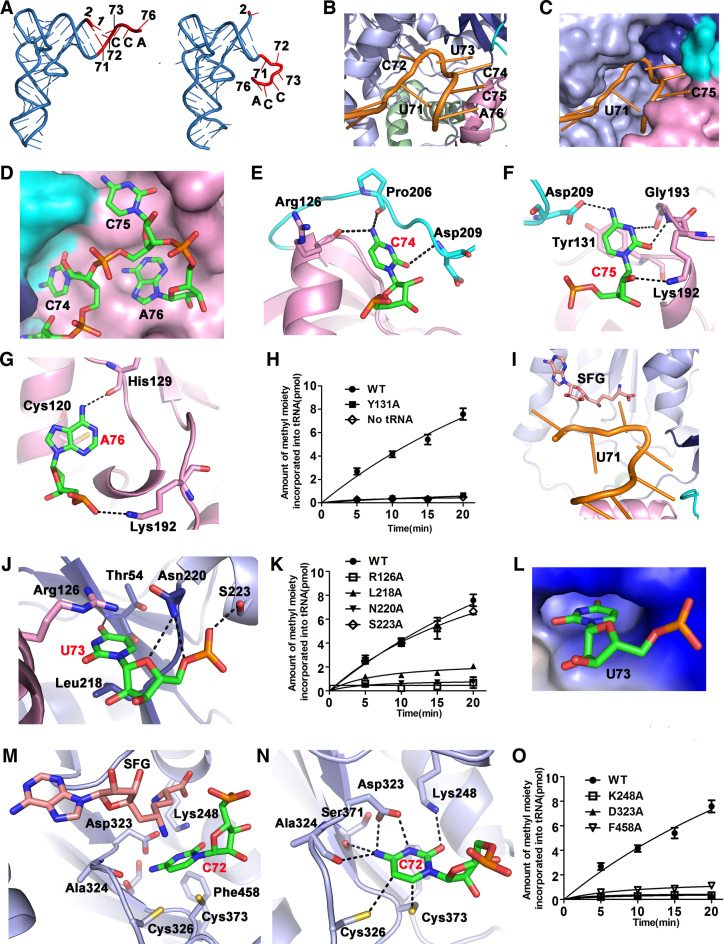
5-methylcytosine (m5C) modifications of RNA are ubiquitous in nature and play important roles in many biological processes such as protein translational regulation, RNA processing and stress response. Aberrant expressions of RNA:m5C methyltransferases are closely associated with various human diseases including cancers. However, no structural information for RNA-bound RNA:m5C methyltransferase was available until now, hindering elucidation of the catalytic mechanism behind RNA:m5C methylation. Here, we have solved the structures of NSun6, a human tRNA:m5C methyltransferase, in the apo form and in complex with a full-length tRNA substrate. These structures show a non-canonical conformation of the bound tRNA, rendering the base moiety of the target cytosine accessible to the enzyme for methylation. Further biochemical assays reveal the critical, but distinct, roles of two conserved cysteine residues for the RNA:m5C methylation. Collectively, for the first time, we have solved the complex structure of a RNA:m5C methyltransferase and addressed the catalytic mechanism of the RNA:m5C methyltransferase family, which may allow for structure-based drug design toward RNA:m5C methyltransferase-related diseases.
© The Author(s) 2017. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures





Similar articles
-
Archaeal NSUN6 catalyzes m5C72 modification on a wide-range of specific tRNAs.Nucleic Acids Res. 2019 Feb 28;47(4):2041-2055. doi: 10.1093/nar/gky1236. Nucleic Acids Res. 2019. PMID: 30541086 Free PMC article.
-
Mechanistic features of the atypical tRNA m1G9 SPOUT methyltransferase, Trm10.Nucleic Acids Res. 2017 Sep 6;45(15):9019-9029. doi: 10.1093/nar/gkx620. Nucleic Acids Res. 2017. PMID: 28911116 Free PMC article.
-
Sequence-structure-function studies of tRNA:m5C methyltransferase Trm4p and its relationship to DNA:m5C and RNA:m5U methyltransferases.Nucleic Acids Res. 2004 Apr 30;32(8):2453-63. doi: 10.1093/nar/gkh564. Print 2004. Nucleic Acids Res. 2004. PMID: 15121902 Free PMC article.
-
Diversity in mechanism and function of tRNA methyltransferases.RNA Biol. 2015;12(4):398-411. doi: 10.1080/15476286.2015.1008358. RNA Biol. 2015. PMID: 25626150 Free PMC article. Review.
-
Transfer RNA methyltransferases with a SpoU-TrmD (SPOUT) fold and their modified nucleosides in tRNA.Biomolecules. 2017 Feb 28;7(1):23. doi: 10.3390/biom7010023. Biomolecules. 2017. PMID: 28264529 Free PMC article. Review.
Cited by
-
Methylation modifications in tRNA and associated disorders: Current research and potential therapeutic targets.Cell Prolif. 2024 Sep;57(9):e13692. doi: 10.1111/cpr.13692. Epub 2024 Jun 28. Cell Prolif. 2024. PMID: 38943267 Free PMC article. Review.
-
Transcriptome-Wide 5-Methylcytosine Functional Profiling of Long Non-Coding RNA in Hepatocellular Carcinoma.Cancer Manag Res. 2020 Aug 4;12:6877-6885. doi: 10.2147/CMAR.S262450. eCollection 2020. Cancer Manag Res. 2020. PMID: 32801911 Free PMC article.
-
Archaeal NSUN6 catalyzes m5C72 modification on a wide-range of specific tRNAs.Nucleic Acids Res. 2019 Feb 28;47(4):2041-2055. doi: 10.1093/nar/gky1236. Nucleic Acids Res. 2019. PMID: 30541086 Free PMC article.
-
Crystal structure of human cytoplasmic tRNAHis-specific 5'-monomethylphosphate capping enzyme.Nucleic Acids Res. 2020 Feb 20;48(3):1572-1582. doi: 10.1093/nar/gkz1216. Nucleic Acids Res. 2020. PMID: 31919512 Free PMC article.
-
A Cross-Tissue Investigation of Molecular Targets and Physiological Functions of Nsun6 Using Knockout Mice.Int J Mol Sci. 2022 Jun 13;23(12):6584. doi: 10.3390/ijms23126584. Int J Mol Sci. 2022. PMID: 35743028 Free PMC article.
References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
