

Selective regulation of Notch ligands during angiogenesis is mediated by vimentin
- PMID: 28533359
- PMCID: PMC5468602
- DOI: 10.1073/pnas.1703057114
Selective regulation of Notch ligands during angiogenesis is mediated by vimentin
Abstract
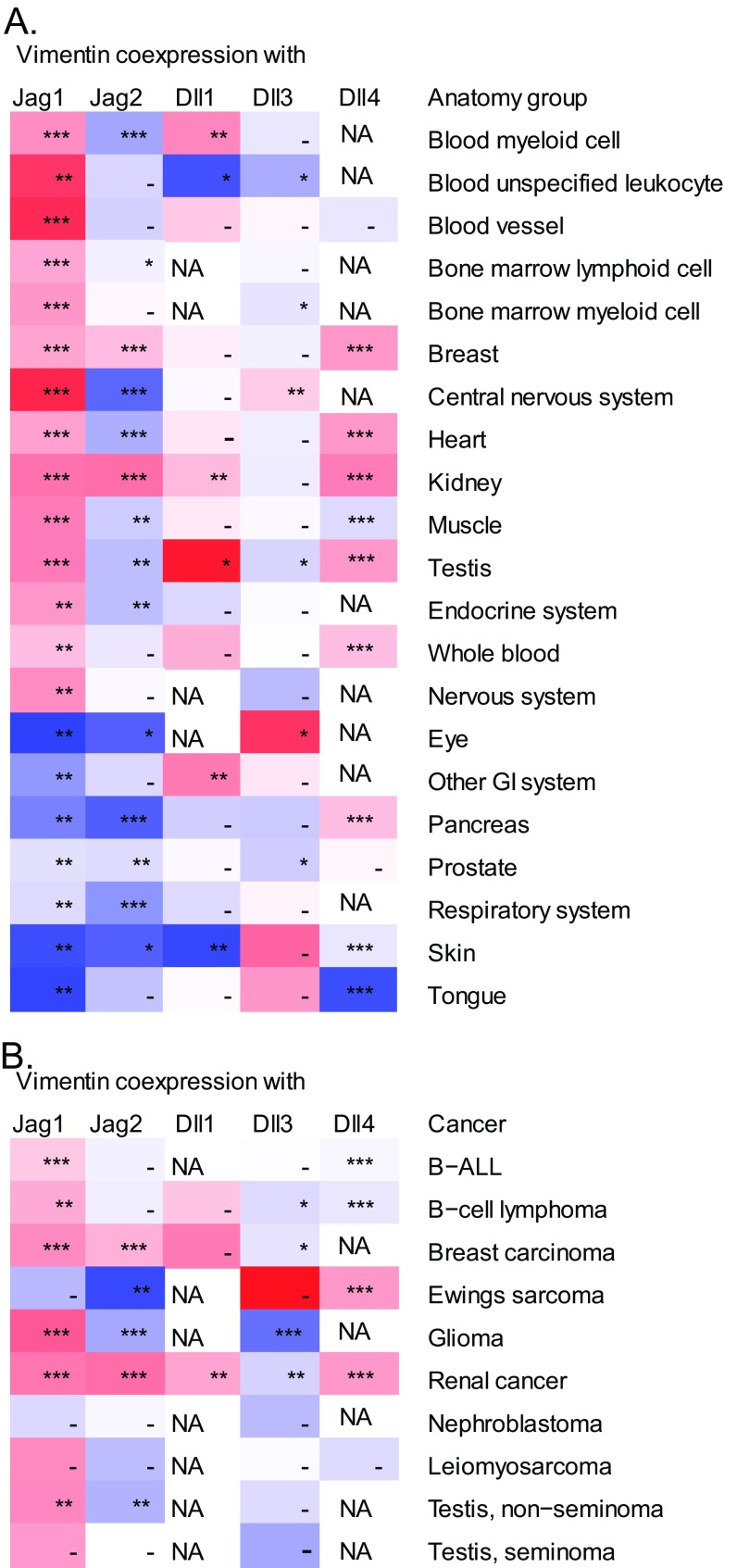
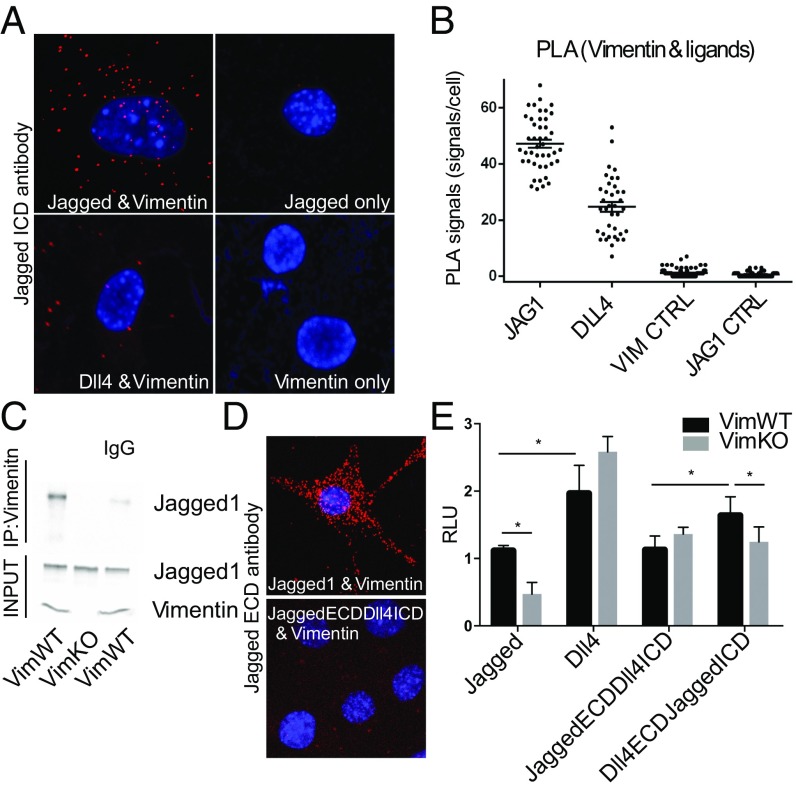
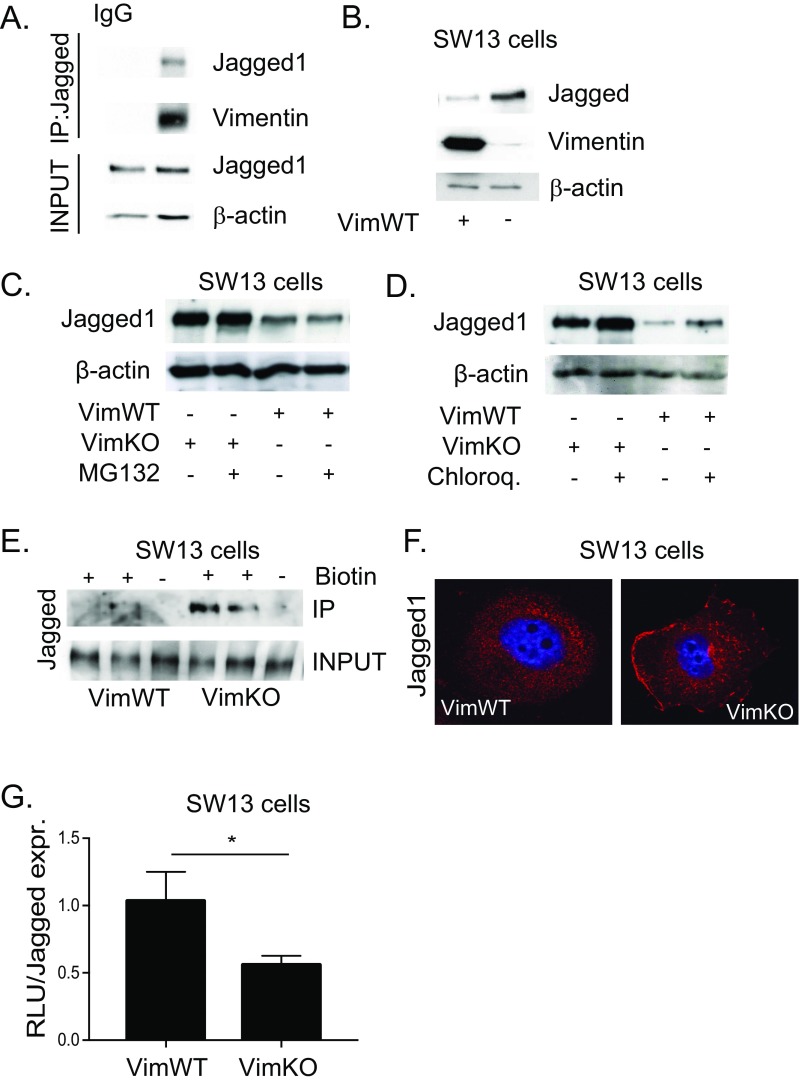
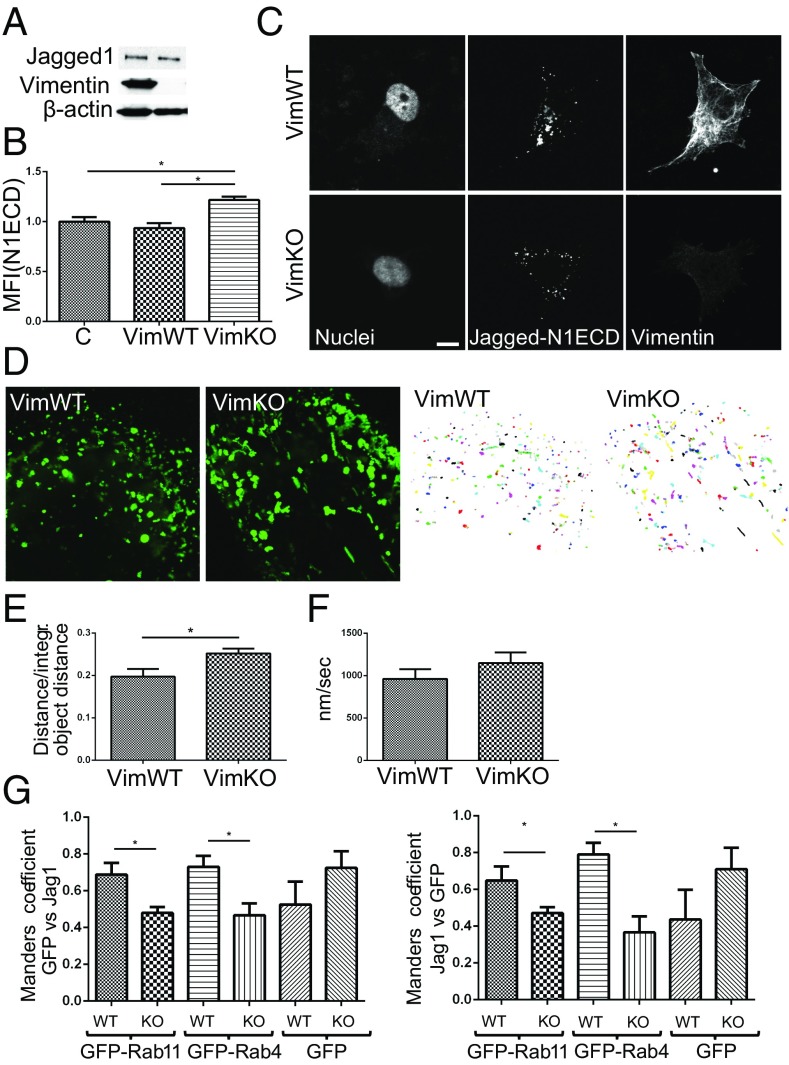
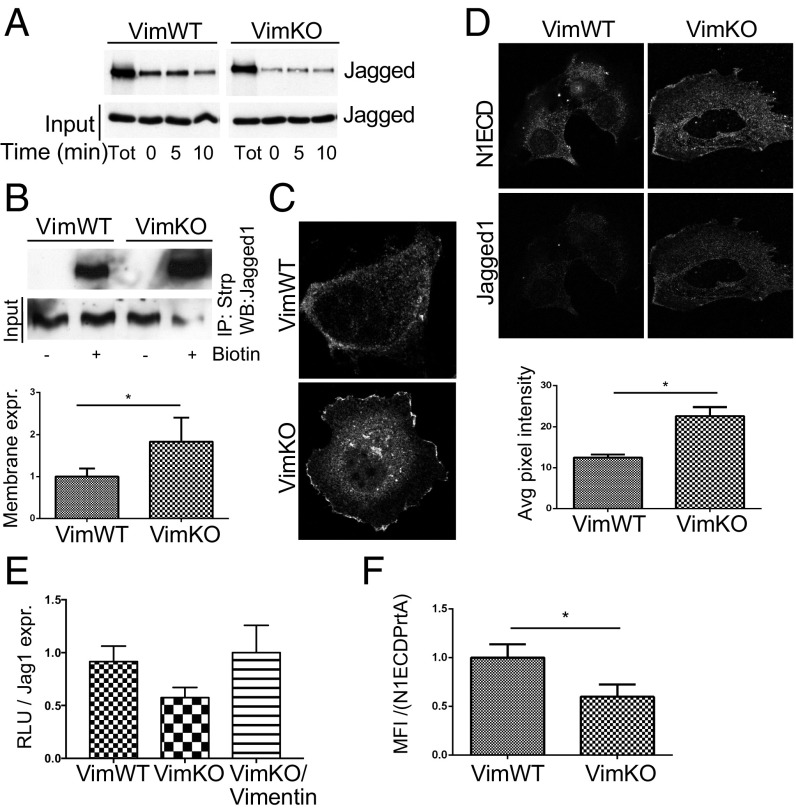
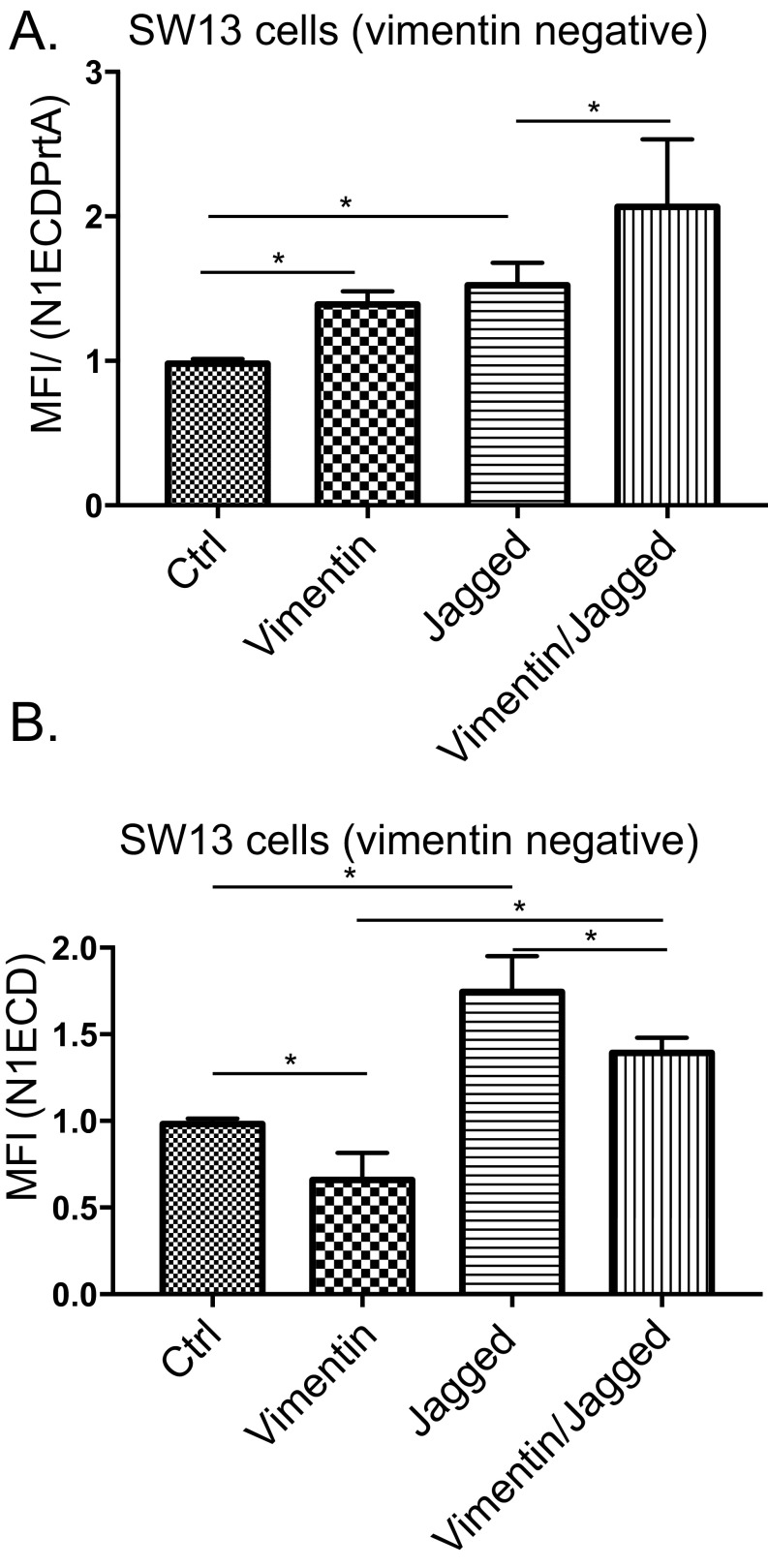
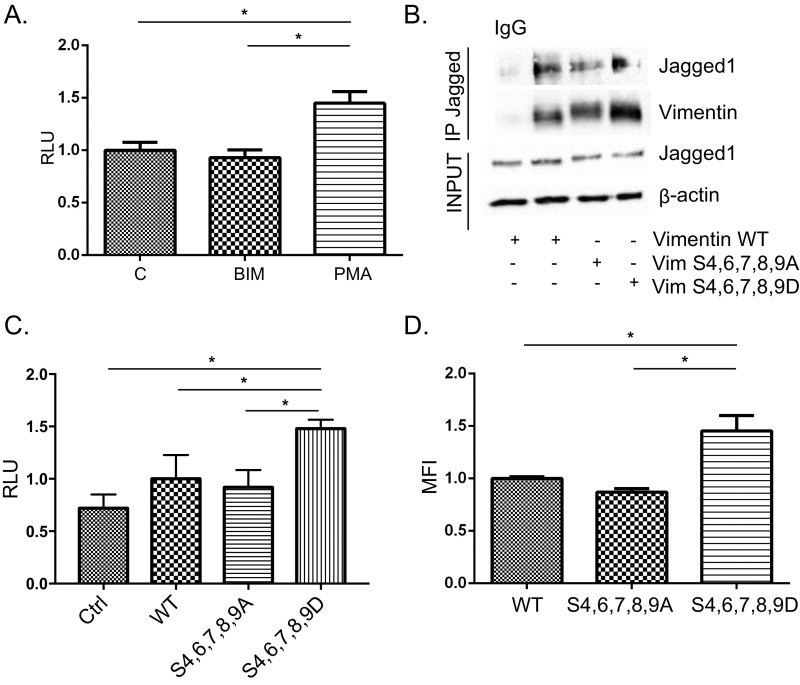
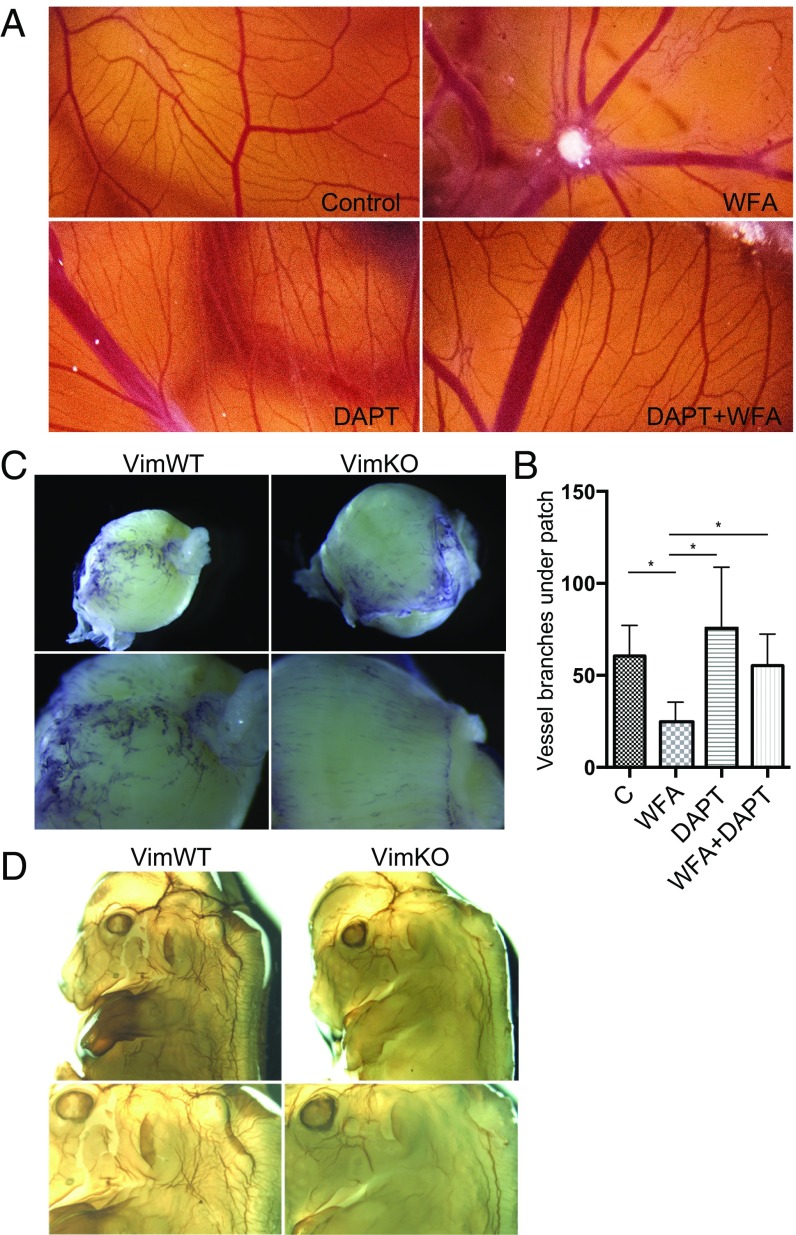
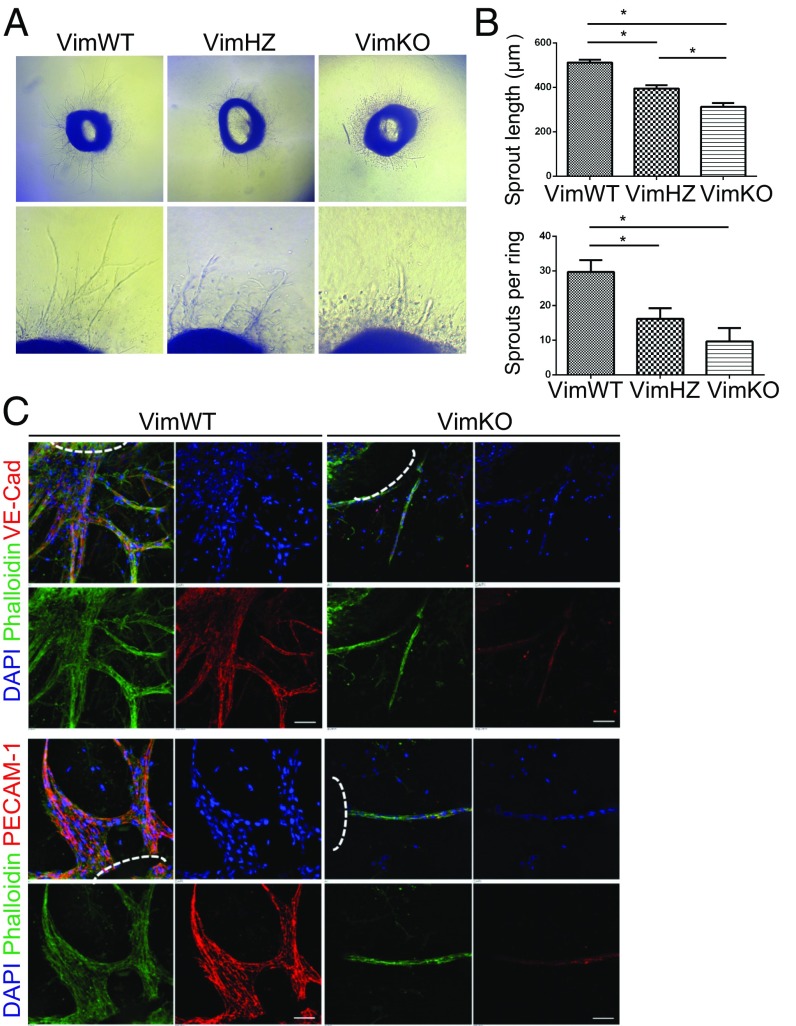
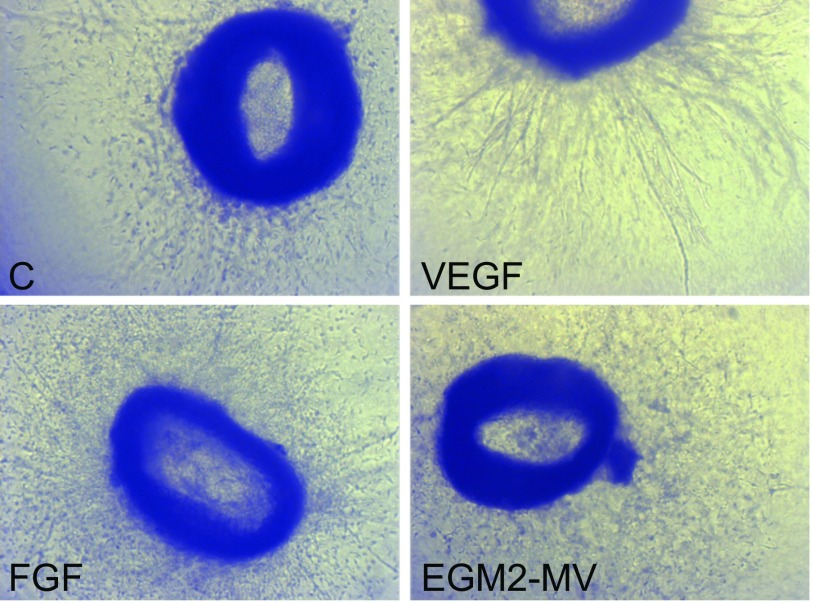
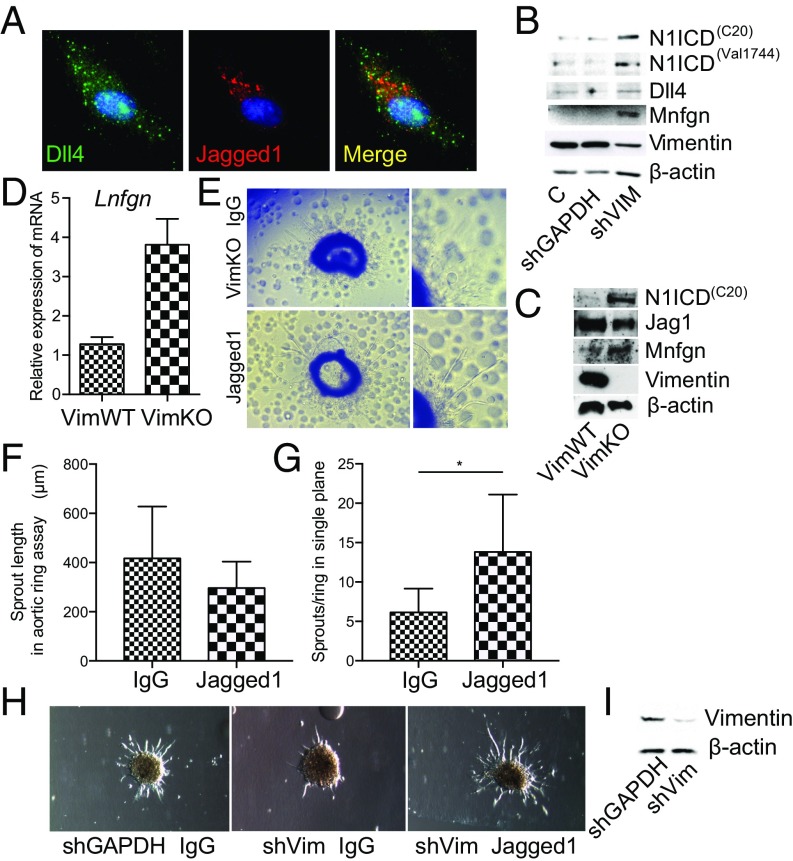
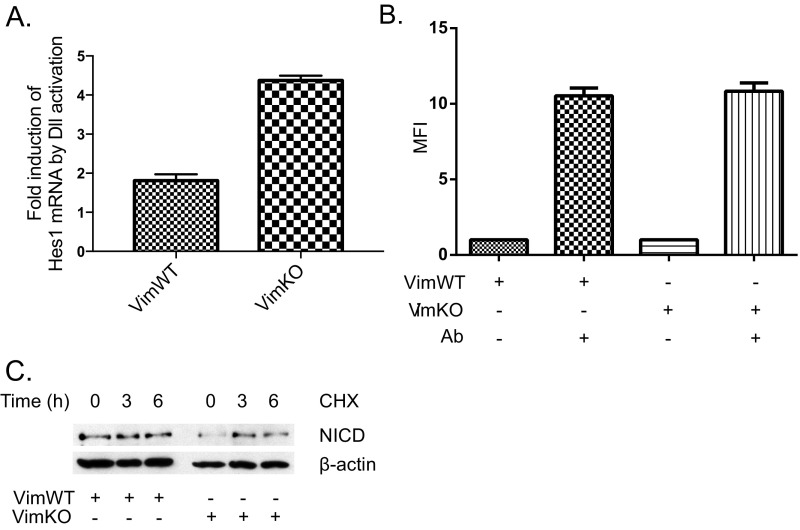
Notch signaling is a key regulator of angiogenesis, in which sprouting is regulated by an equilibrium between inhibitory Dll4-Notch signaling and promoting Jagged-Notch signaling. Whereas Fringe proteins modify Notch receptors and strengthen their activation by Dll4 ligands, other mechanisms balancing Jagged and Dll4 signaling are yet to be described. The intermediate filament protein vimentin, which has been previously shown to affect vascular integrity and regenerative signaling, is here shown to regulate ligand-specific Notch signaling. Vimentin interacts with Jagged, impedes basal recycling endocytosis of ligands, but is required for efficient receptor ligand transendocytosis and Notch activation upon receptor binding. Analyses of Notch signal activation by using chimeric ligands with swapped intracellular domains (ICDs), demonstrated that the Jagged ICD binds to vimentin and contributes to signaling strength. Vimentin also suppresses expression of Fringe proteins, whereas depletion of vimentin enhances Fringe levels to promote Dll4 signaling. In line with these data, the vasculature in vimentin knockout (VimKO) embryos and placental tissue is underdeveloped with reduced branching. Disrupted angiogenesis in aortic rings from VimKO mice and in endothelial 3D sprouting assays can be rescued by reactivating Notch signaling by recombinant Jagged ligands. Taken together, we reveal a function of vimentin and demonstrate that vimentin regulates Notch ligand signaling activities during angiogenesis.
Keywords: Jagged; Notch; angiogenesis; vimentin.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Schiffers PM, et al. Altered flow-induced arterial remodeling in vimentin-deficient mice. Arterioscler Thromb Vasc Biol. 2000;20:611–616. - PubMed
-
- Nieminen M, et al. Vimentin function in lymphocyte adhesion and transcellular migration. Nat Cell Biol. 2006;8:156–162. - PubMed
-
- Pallari HM, Eriksson JE. Intermediate filaments as signaling platforms. Sci STKE. 2006;2006:pe53. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
