

Mechanosensing of shear by Pseudomonas aeruginosa leads to increased levels of the cyclic-di-GMP signal initiating biofilm development
- PMID: 28533383
- PMCID: PMC5468607
- DOI: 10.1073/pnas.1703255114
Mechanosensing of shear by Pseudomonas aeruginosa leads to increased levels of the cyclic-di-GMP signal initiating biofilm development
Erratum in
-
Correction for Rodesney et al., Mechanosensing of shear by Pseudomonas aeruginosa leads to increased levels of the cyclic-di-GMP signal initiating biofilm development.Proc Natl Acad Sci U S A. 2017 Jul 11;114(28):E5760. doi: 10.1073/pnas.1710411114. Epub 2017 Jul 3. Proc Natl Acad Sci U S A. 2017. PMID: 28673990 Free PMC article. No abstract available.
Abstract
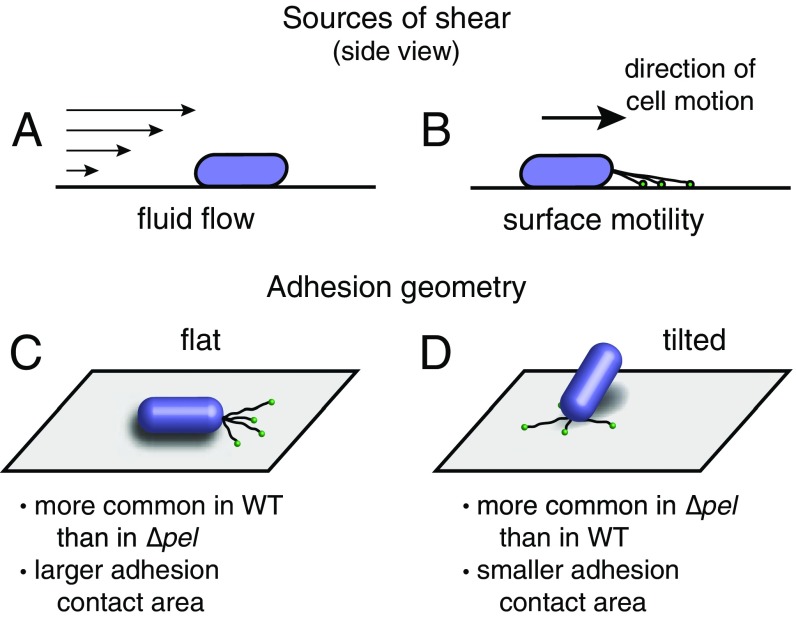
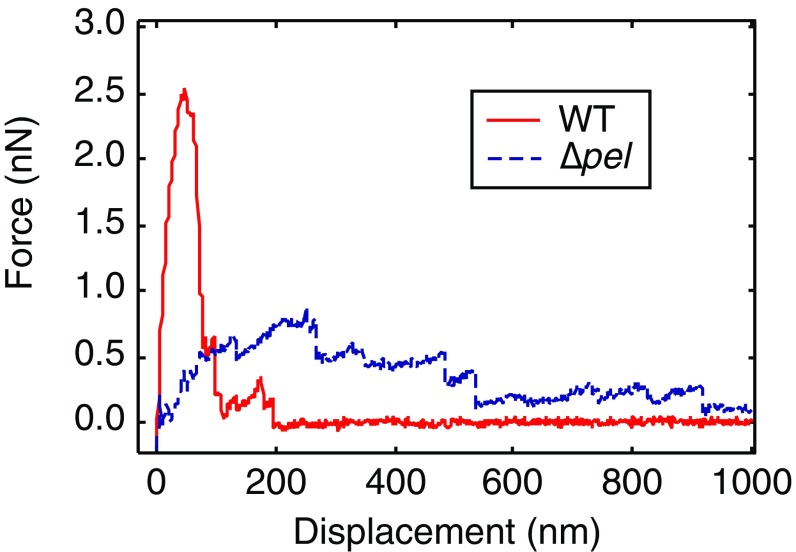
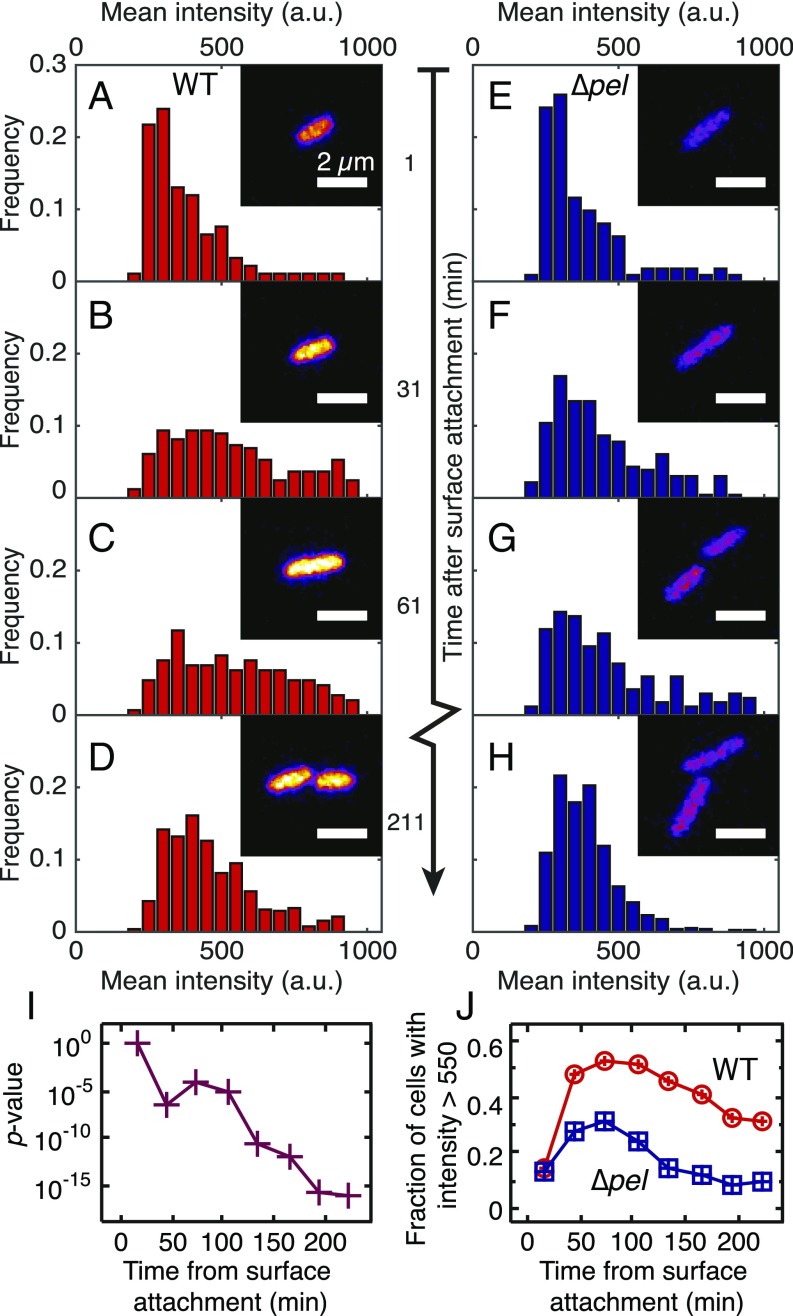
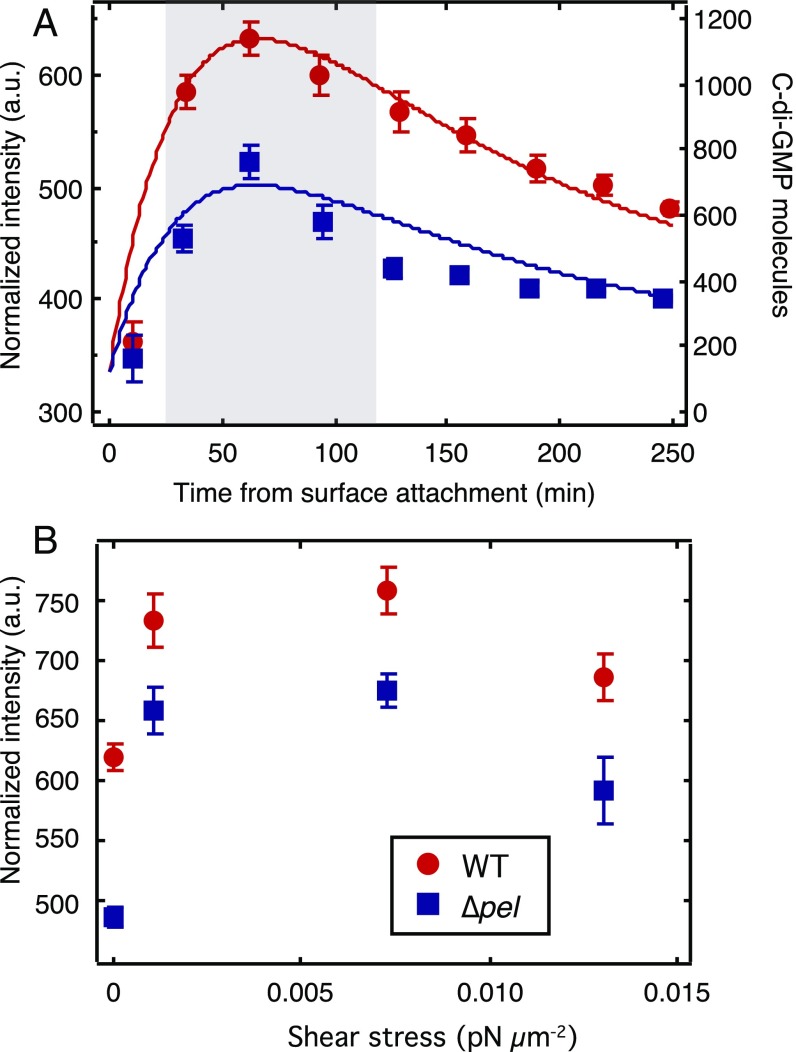
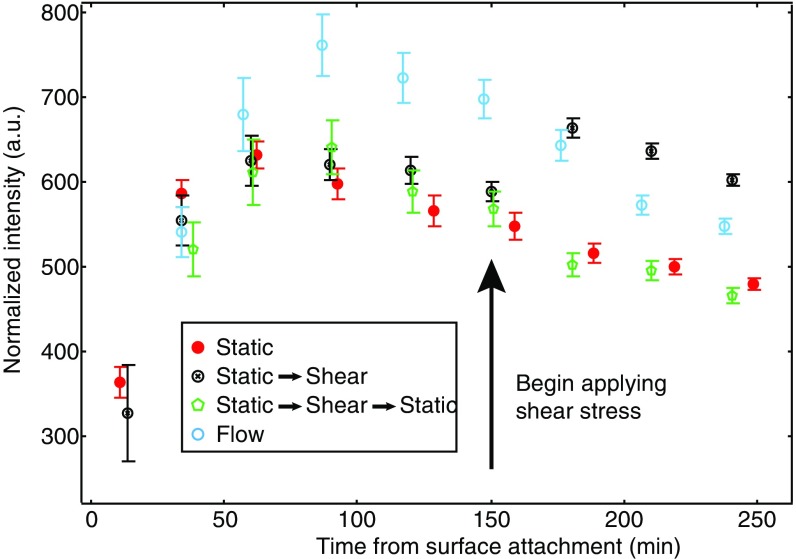
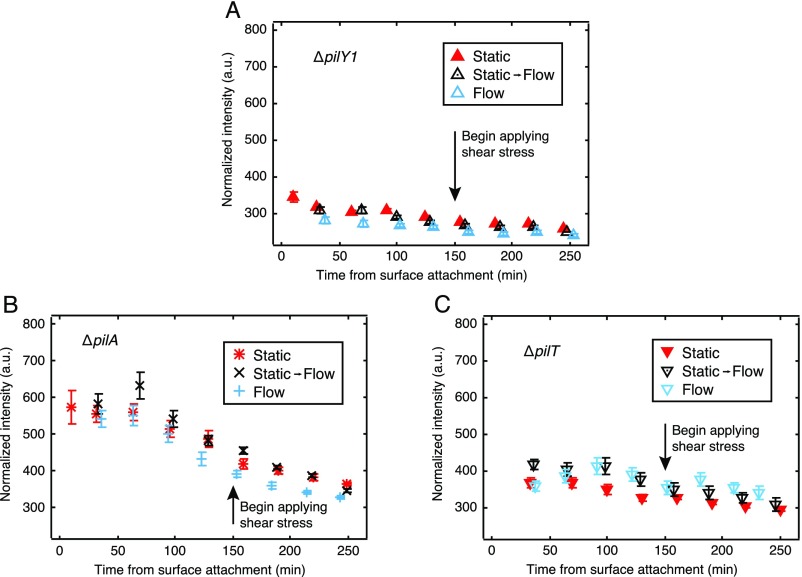
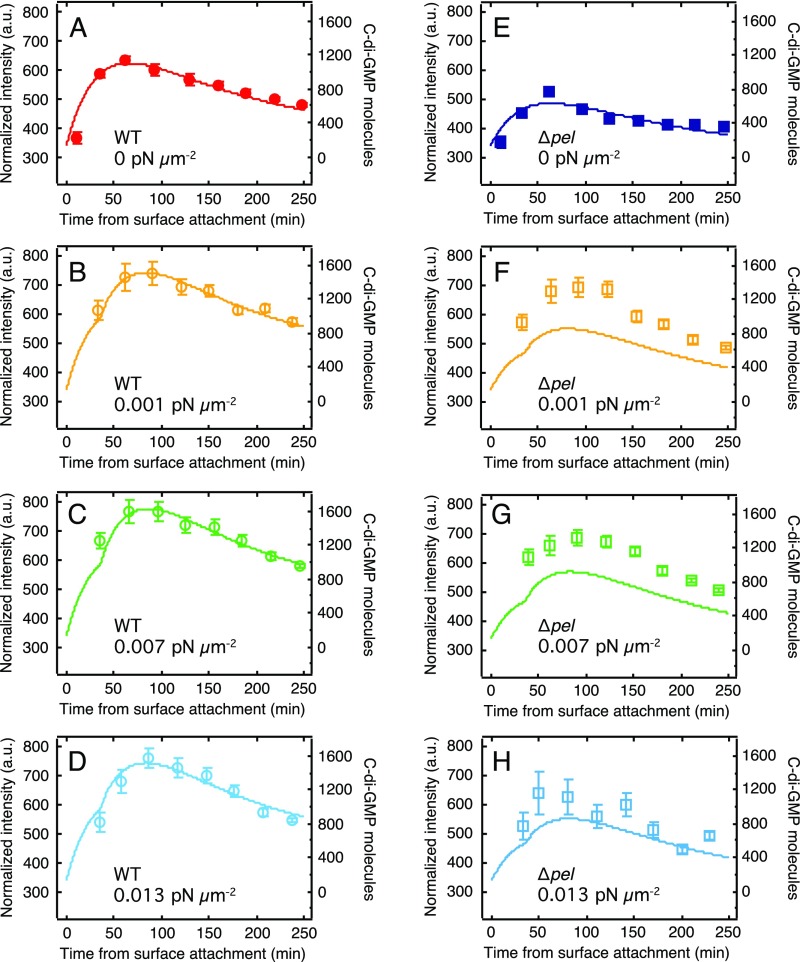
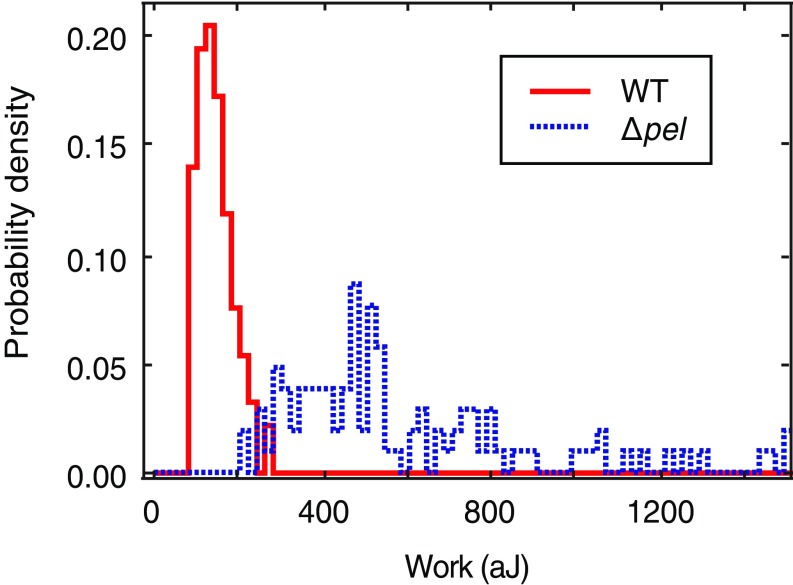
Biofilms are communities of sessile microbes that are phenotypically distinct from their genetically identical, free-swimming counterparts. Biofilms initiate when bacteria attach to a solid surface. Attachment triggers intracellular signaling to change gene expression from the planktonic to the biofilm phenotype. For Pseudomonas aeruginosa, it has long been known that intracellular levels of the signal cyclic-di-GMP increase upon surface adhesion and that this is required to begin biofilm development. However, what cue is sensed to notify bacteria that they are attached to the surface has not been known. Here, we show that mechanical shear acts as a cue for surface adhesion and activates cyclic-di-GMP signaling. The magnitude of the shear force, and thereby the corresponding activation of cyclic-di-GMP signaling, can be adjusted both by varying the strength of the adhesion that binds bacteria to the surface and by varying the rate of fluid flow over surface-bound bacteria. We show that the envelope protein PilY1 and functional type IV pili are required mechanosensory elements. An analytic model that accounts for the feedback between mechanosensors, cyclic-di-GMP signaling, and production of adhesive polysaccharides describes our data well.
Keywords: Pseudomonas aeruginosa; biofilm; cyclic-di-GMP; mechanobiology; mechanosensing.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
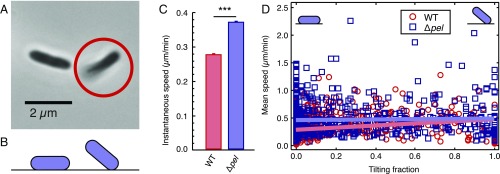
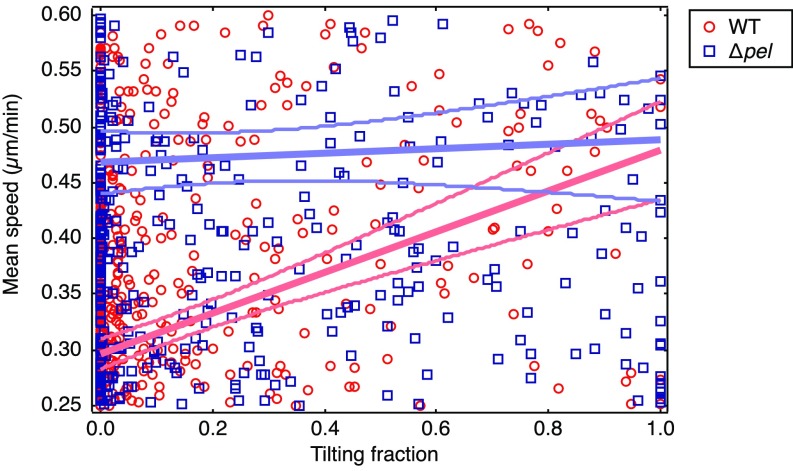
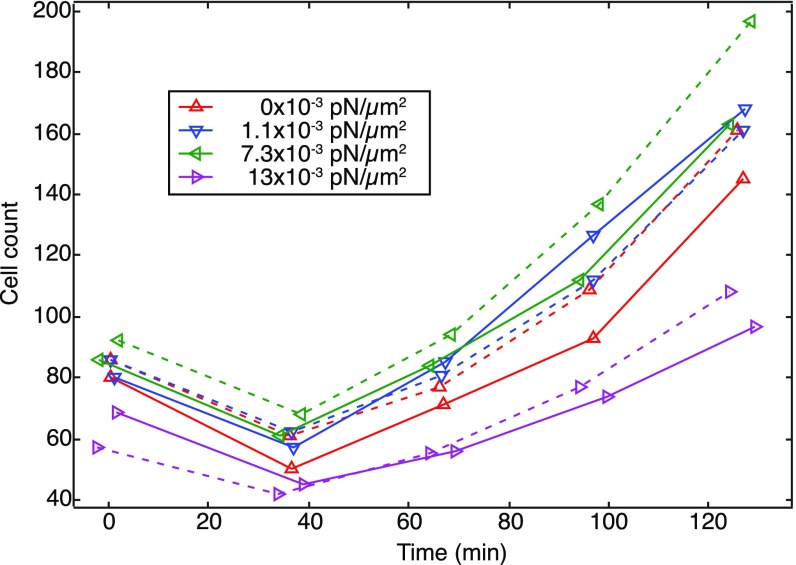
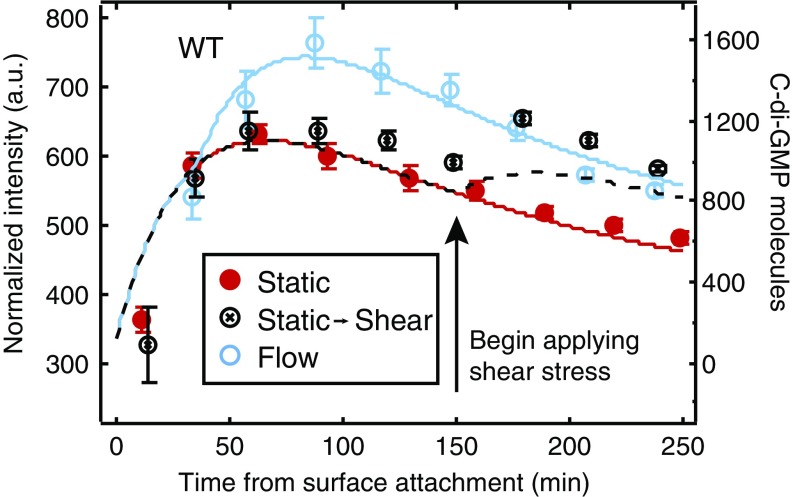
References
-
- Wolcott R, Dowd S. The role of biofilms: Are we hitting the right target? Plast Reconstr Surg. 2011;127:28S–35S. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
