

Selective lowering of synapsins induced by oligomeric α-synuclein exacerbates memory deficits
- PMID: 28533388
- PMCID: PMC5468636
- DOI: 10.1073/pnas.1704698114
Selective lowering of synapsins induced by oligomeric α-synuclein exacerbates memory deficits
Abstract
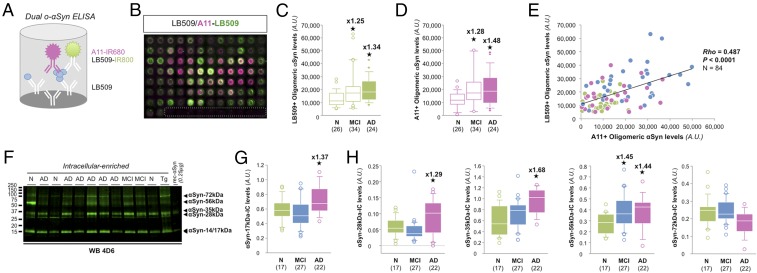
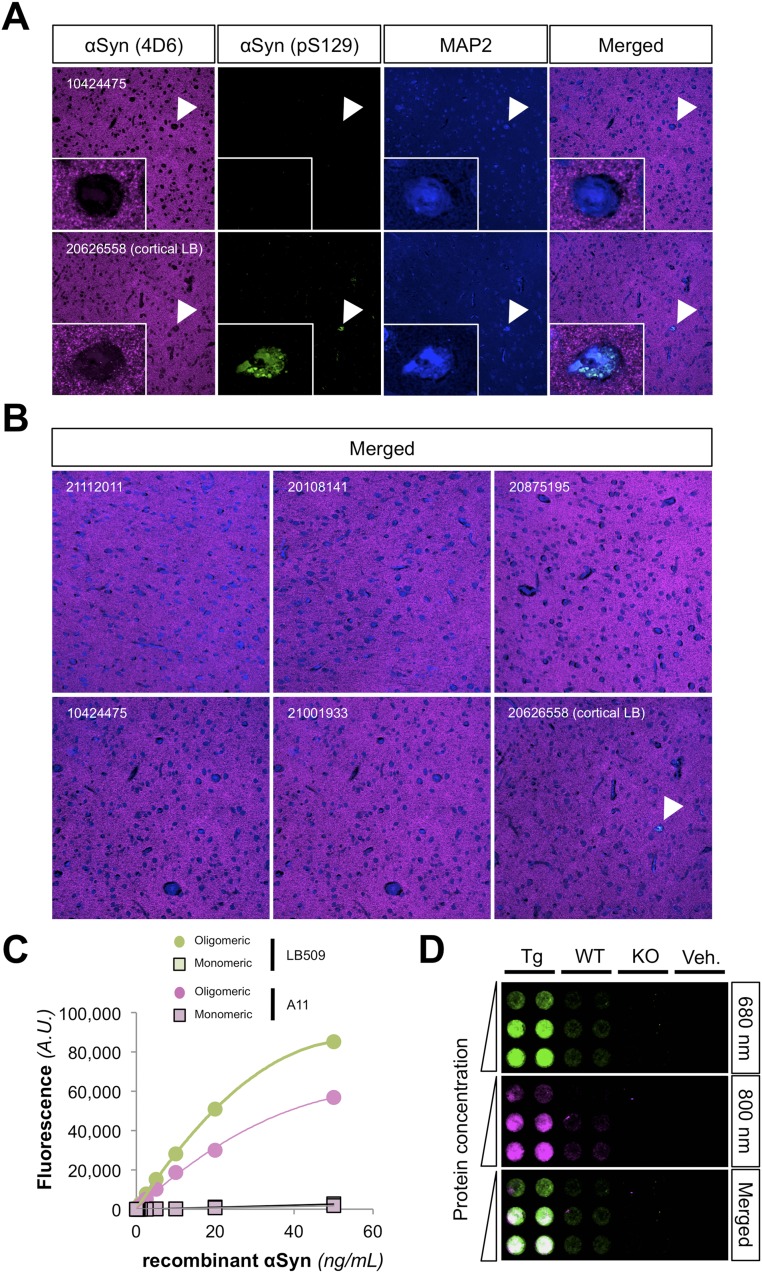
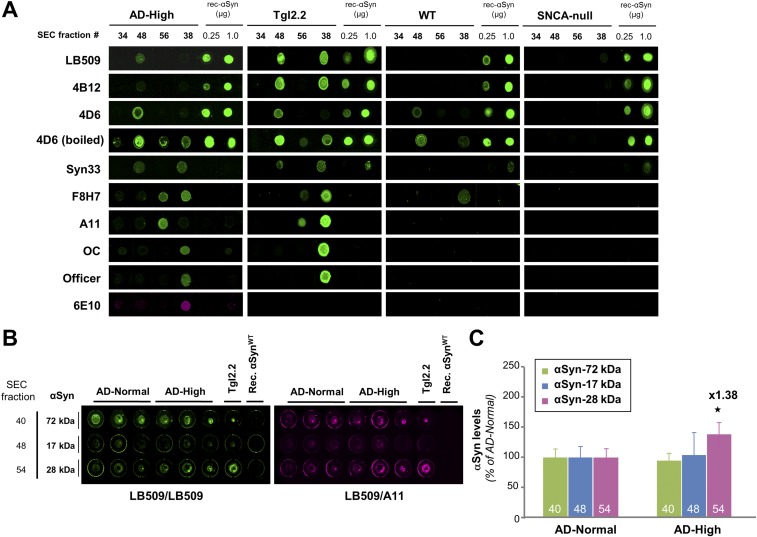
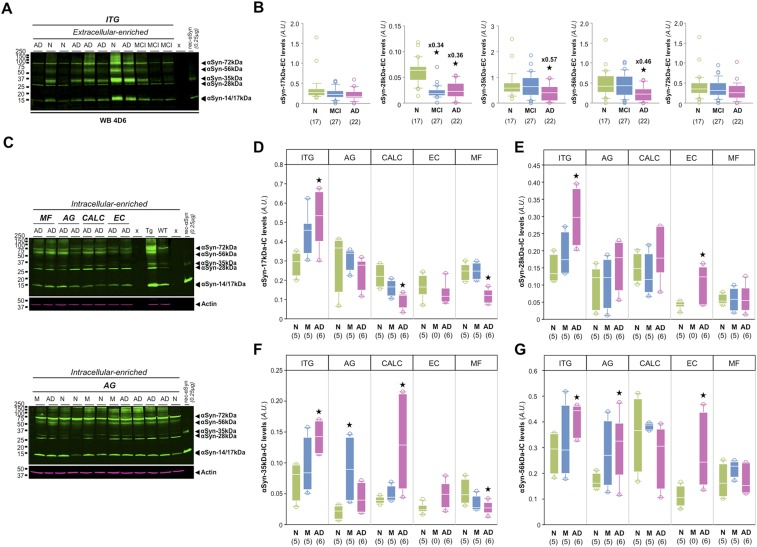
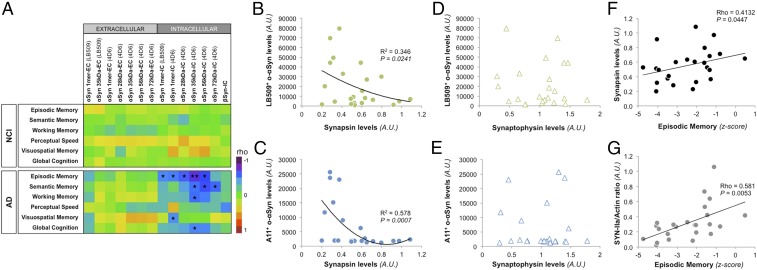
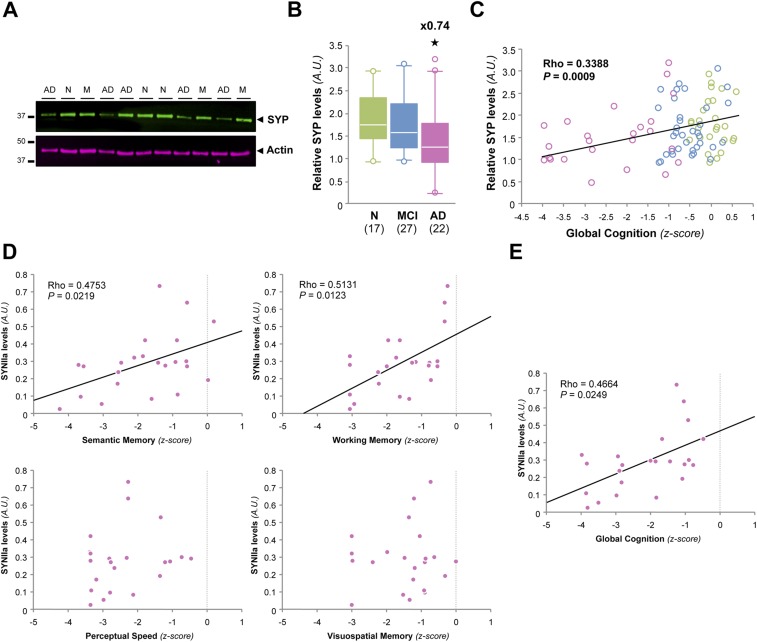
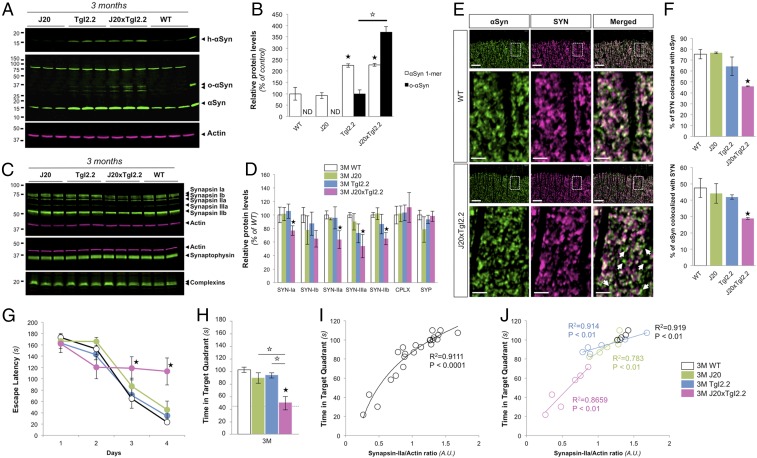
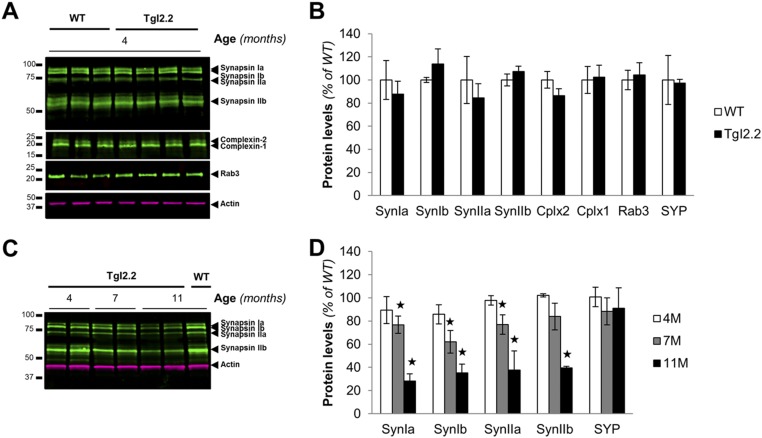
Mounting evidence indicates that soluble oligomeric forms of amyloid proteins linked to neurodegenerative disorders, such as amyloid-β (Aβ), tau, or α-synuclein (αSyn) might be the major deleterious species for neuronal function in these diseases. Here, we found an abnormal accumulation of oligomeric αSyn species in AD brains by custom ELISA, size-exclusion chromatography, and nondenaturing/denaturing immunoblotting techniques. Importantly, the abundance of αSyn oligomers in human brain tissue correlated with cognitive impairment and reductions in synapsin expression. By overexpressing WT human αSyn in an AD mouse model, we artificially enhanced αSyn oligomerization. These bigenic mice displayed exacerbated Aβ-induced cognitive deficits and a selective decrease in synapsins. Following isolation of various soluble αSyn assemblies from transgenic mice, we found that in vitro delivery of exogenous oligomeric αSyn but not monomeric αSyn was causing a lowering in synapsin-I/II protein abundance. For a particular αSyn oligomer, these changes were either dependent or independent on endogenous αSyn expression. Finally, at a molecular level, the expression of synapsin genes SYN1 and SYN2 was down-regulated in vivo and in vitro by αSyn oligomers, which decreased two transcription factors, cAMP response element binding and Nurr1, controlling synapsin gene promoter activity. Overall, our results demonstrate that endogenous αSyn oligomers can impair memory by selectively lowering synapsin expression.
Keywords: Alzheimer’s disease; memory; oligomer; synapsins; α-synuclein.
Conflict of interest statement
The authors declare no conflict of interest.
Figures




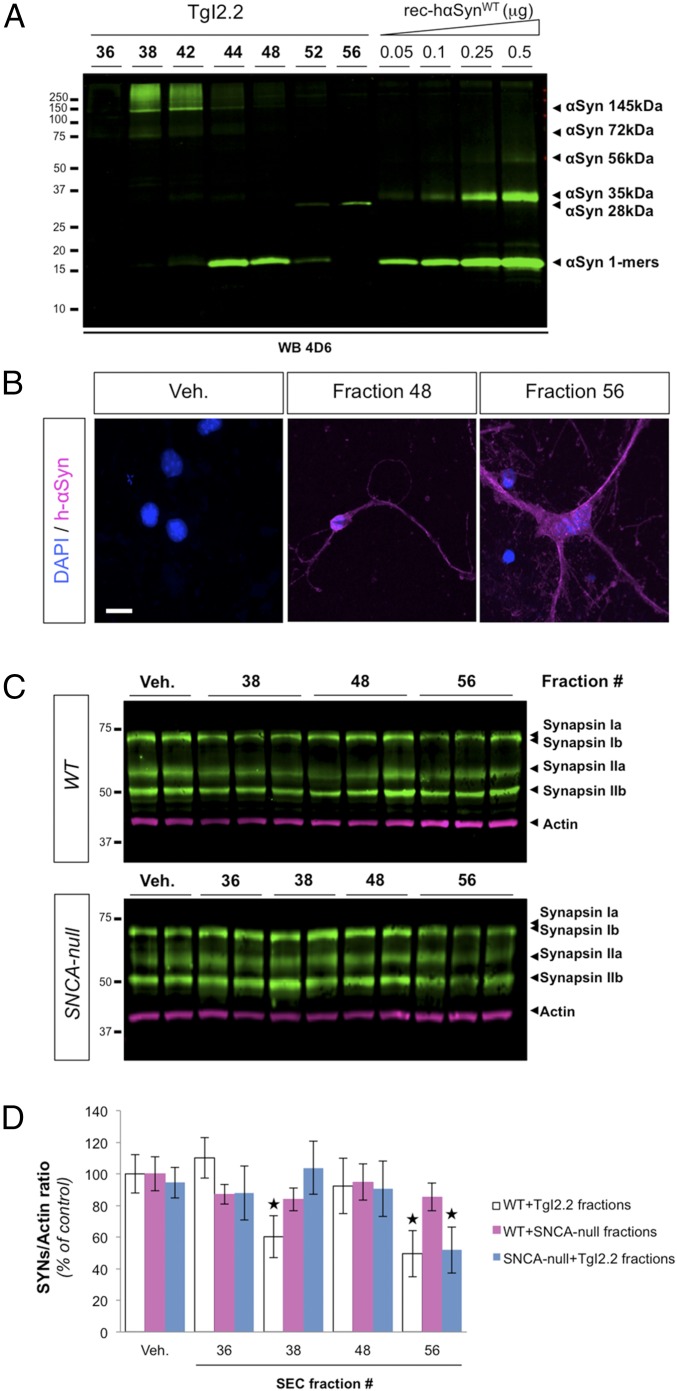
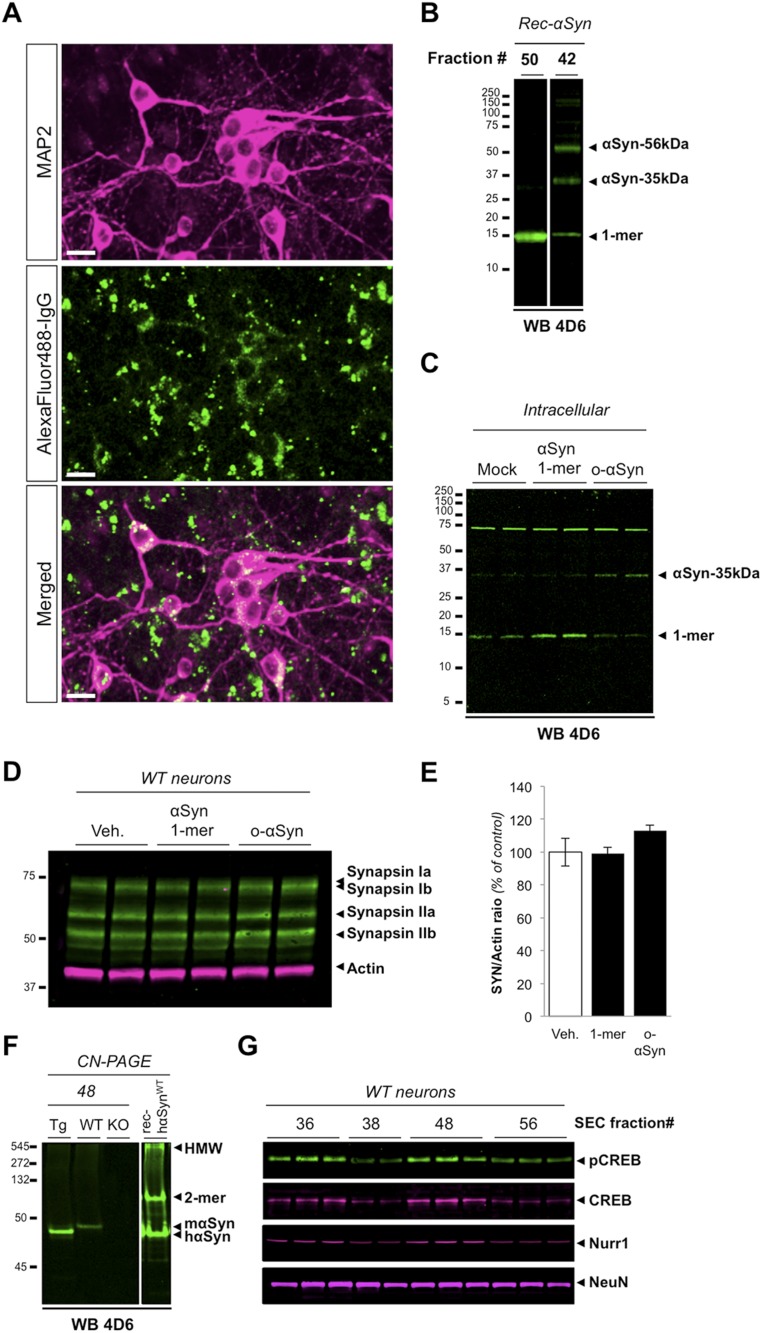
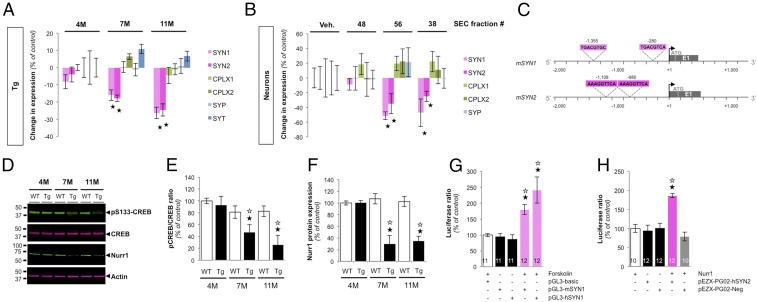
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
