

Angular measurements of the dynein ring reveal a stepping mechanism dependent on a flexible stalk
- PMID: 28533393
- PMCID: PMC5468668
- DOI: 10.1073/pnas.1620149114
Angular measurements of the dynein ring reveal a stepping mechanism dependent on a flexible stalk
Abstract
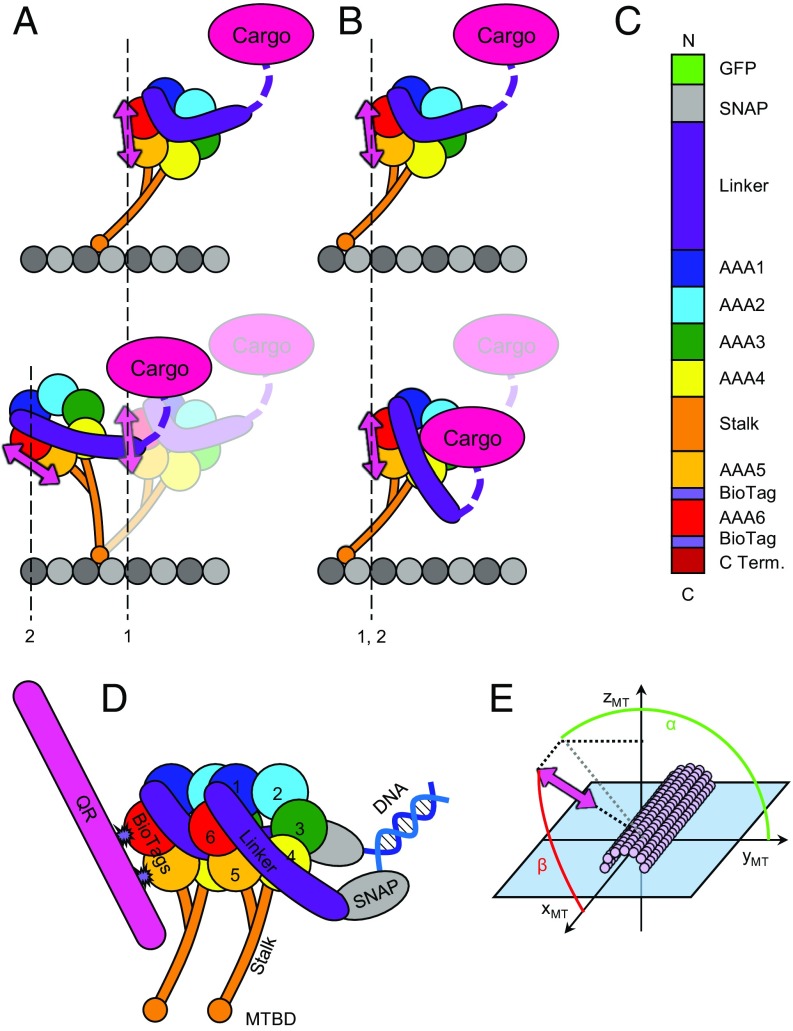
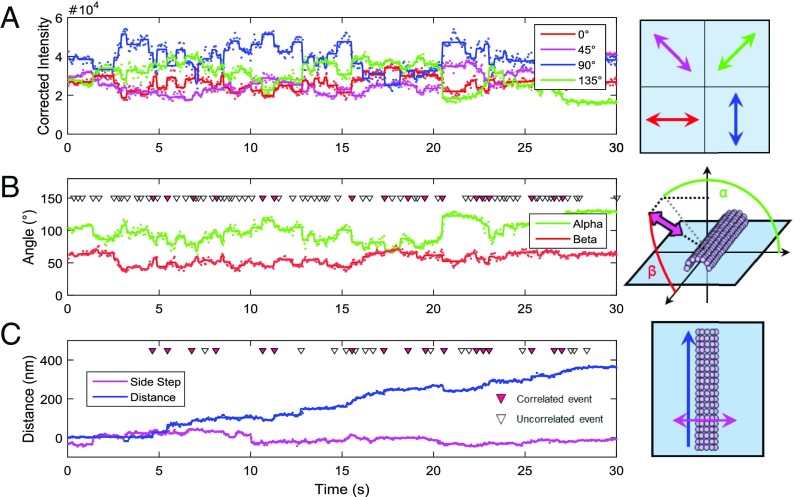
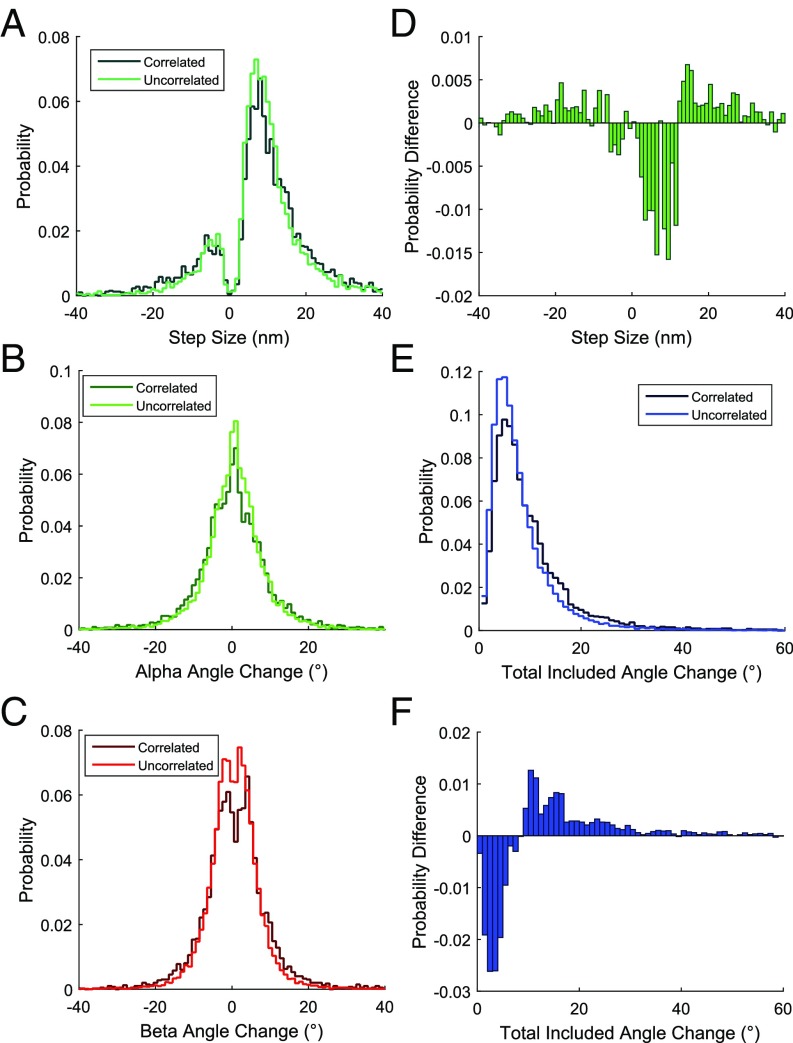
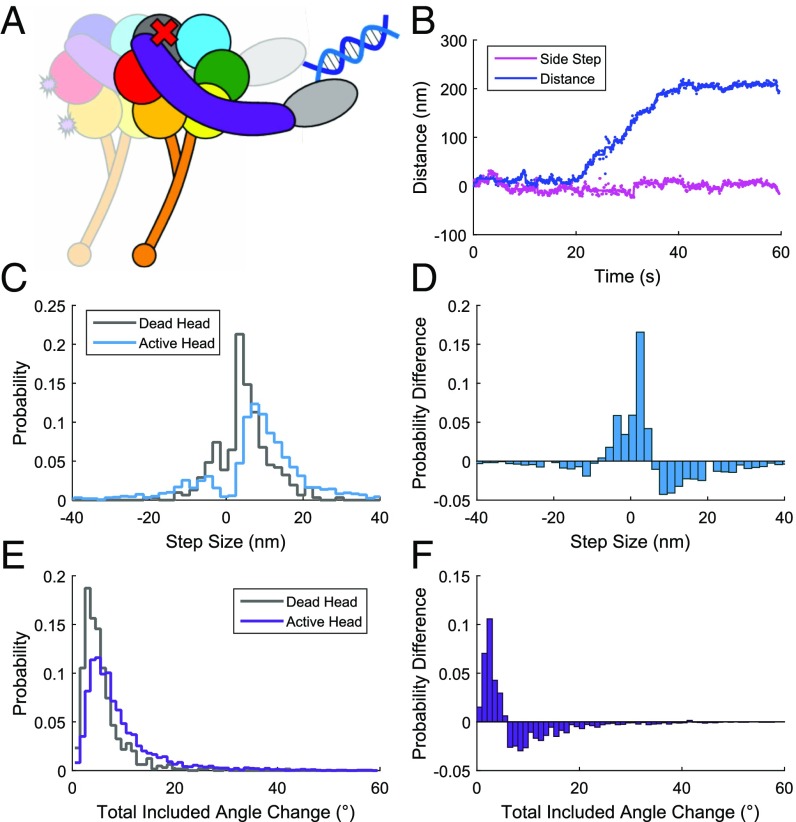
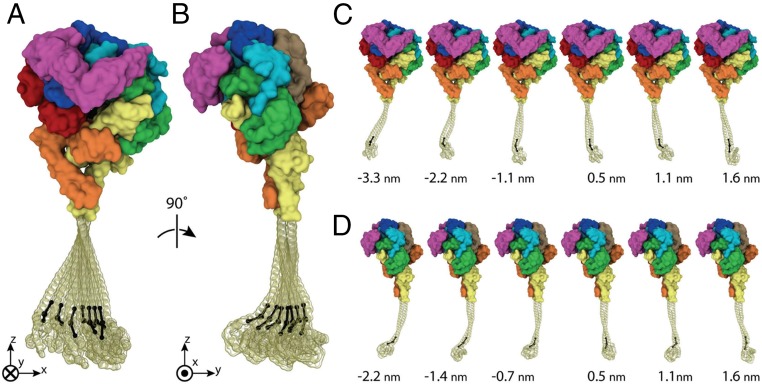
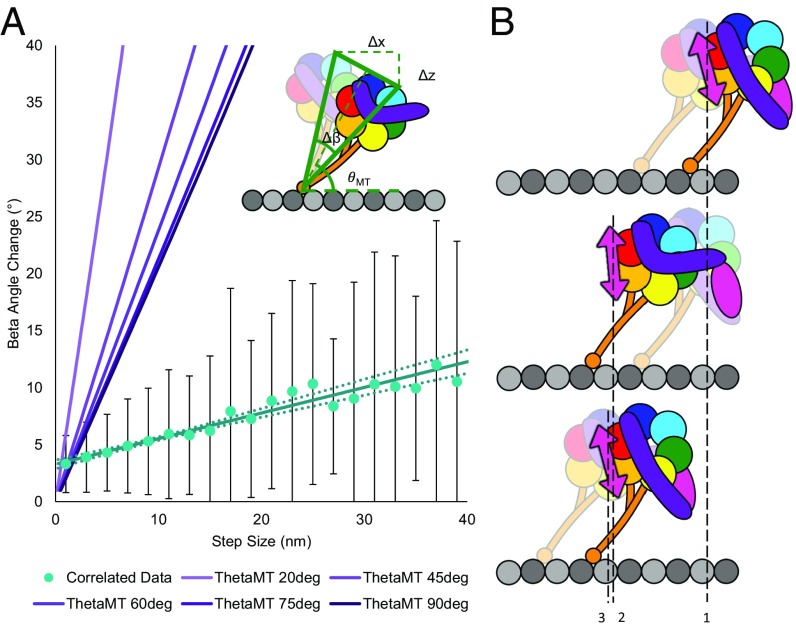
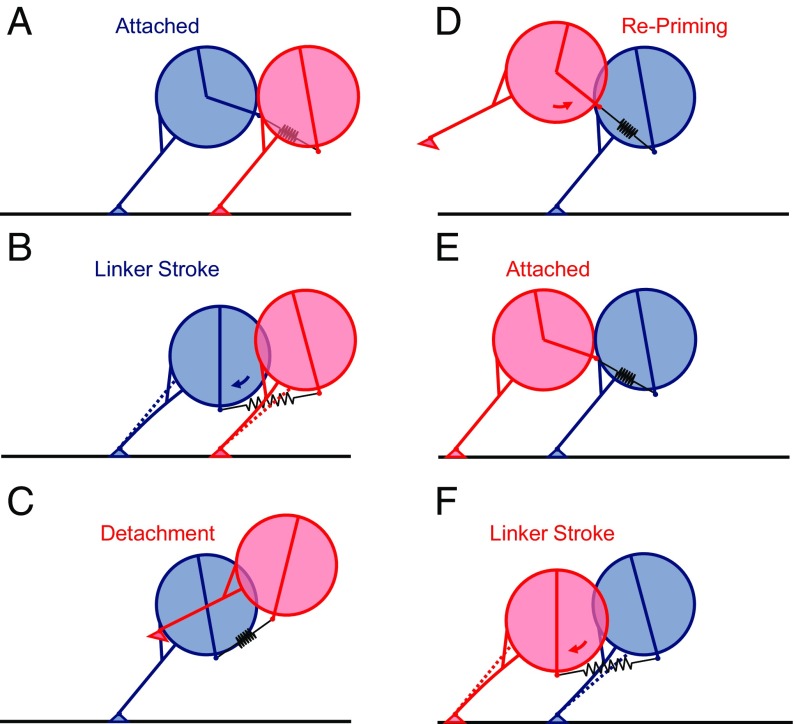
The force-generating mechanism of dynein differs from the force-generating mechanisms of other cytoskeletal motors. To examine the structural dynamics of dynein's stepping mechanism in real time, we used polarized total internal reflection fluorescence microscopy with nanometer accuracy localization to track the orientation and position of single motors. By measuring the polarized emission of individual quantum nanorods coupled to the dynein ring, we determined the angular position of the ring and found that it rotates relative to the microtubule (MT) while walking. Surprisingly, the observed rotations were small, averaging only 8.3°, and were only weakly correlated with steps. Measurements at two independent labeling positions on opposite sides of the ring showed similar small rotations. Our results are inconsistent with a classic power-stroke mechanism, and instead support a flexible stalk model in which interhead strain rotates the rings through bending and hinging of the stalk. Mechanical compliances of the stalk and hinge determined based on a 3.3-μs molecular dynamics simulation account for the degree of ring rotation observed experimentally. Together, these observations demonstrate that the stepping mechanism of dynein is fundamentally different from the stepping mechanisms of other well-studied MT motors, because it is characterized by constant small-scale fluctuations of a large but flexible structure fully consistent with the variable stepping pattern observed as dynein moves along the MT.
Keywords: TIRF; dynein; molecular dynamics; polarization; single molecule.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Direct observation shows superposition and large scale flexibility within cytoplasmic dynein motors moving along microtubules.Nat Commun. 2015 Sep 14;6:8179. doi: 10.1038/ncomms9179. Nat Commun. 2015. PMID: 26365535 Free PMC article.
-
Modeling of Chemomechanical Coupling of Cytoplasmic Dynein Motors.J Phys Chem B. 2024 Oct 17;128(41):10063-10074. doi: 10.1021/acs.jpcb.4c04554. Epub 2024 Oct 9. J Phys Chem B. 2024. PMID: 39382058
-
Structural basis for microtubule binding and release by dynein.Science. 2012 Sep 21;337(6101):1532-1536. doi: 10.1126/science.1224151. Science. 2012. PMID: 22997337 Free PMC article.
-
Regulation of processive motion and microtubule localization of cytoplasmic dynein.Biochem Soc Trans. 2015 Feb;43(1):48-57. doi: 10.1042/BST20140252. Biochem Soc Trans. 2015. PMID: 25619245 Review.
-
Molecular organization and force-generating mechanism of dynein.FEBS J. 2011 Sep;278(17):2964-79. doi: 10.1111/j.1742-4658.2011.08253.x. Epub 2011 Aug 8. FEBS J. 2011. PMID: 21777385 Review.
Cited by
-
Fluorescence microscopy applied to intracellular transport by microtubule motors.J Biosci. 2018 Jul;43(3):437-445. J Biosci. 2018. PMID: 30002263 Review.
-
Fundamental Limits on Measuring the Rotational Constraint of Single Molecules Using Fluorescence Microscopy.Phys Rev Lett. 2019 May 17;122(19):198301. doi: 10.1103/PhysRevLett.122.198301. Phys Rev Lett. 2019. PMID: 31144939 Free PMC article.
-
Single-molecule orientation localization microscopy for resolving structural heterogeneities between amyloid fibrils.Optica. 2020 Jun 20;7(6):602-607. doi: 10.1364/optica.388157. Epub 2020 Jun 4. Optica. 2020. PMID: 32832582 Free PMC article.
-
Structure of the Flight Muscle Thick Filament from the Bumble Bee, Bombus ignitus, at 6 Å Resolution.Int J Mol Sci. 2022 Dec 26;24(1):377. doi: 10.3390/ijms24010377. Int J Mol Sci. 2022. PMID: 36613818 Free PMC article.
-
Regulatory mechanisms of the dynein-2 motility by post-translational modification revealed by MD simulation.Sci Rep. 2023 Jan 26;13(1):1477. doi: 10.1038/s41598-023-28026-z. Sci Rep. 2023. PMID: 36702893 Free PMC article.
References
-
- Paschal BM, Vallee RB. Retrograde transport by the microtubule-associated protein MAP 1C. Nature. 1987;330:181–183. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
