

Jasmonoyl-L-Tryptophan Disrupts IAA Activity through the AUX1 Auxin Permease
- PMID: 28533791
- PMCID: PMC5420569
- DOI: 10.3389/fpls.2017.00736
Jasmonoyl-L-Tryptophan Disrupts IAA Activity through the AUX1 Auxin Permease
Abstract
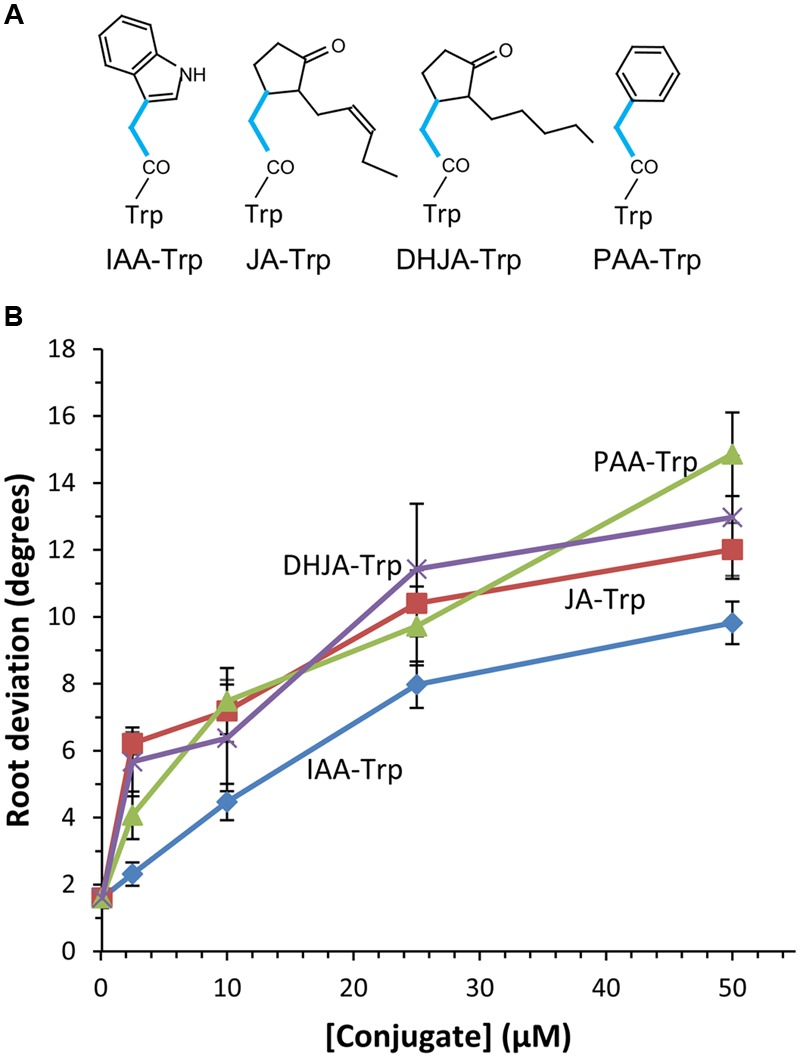
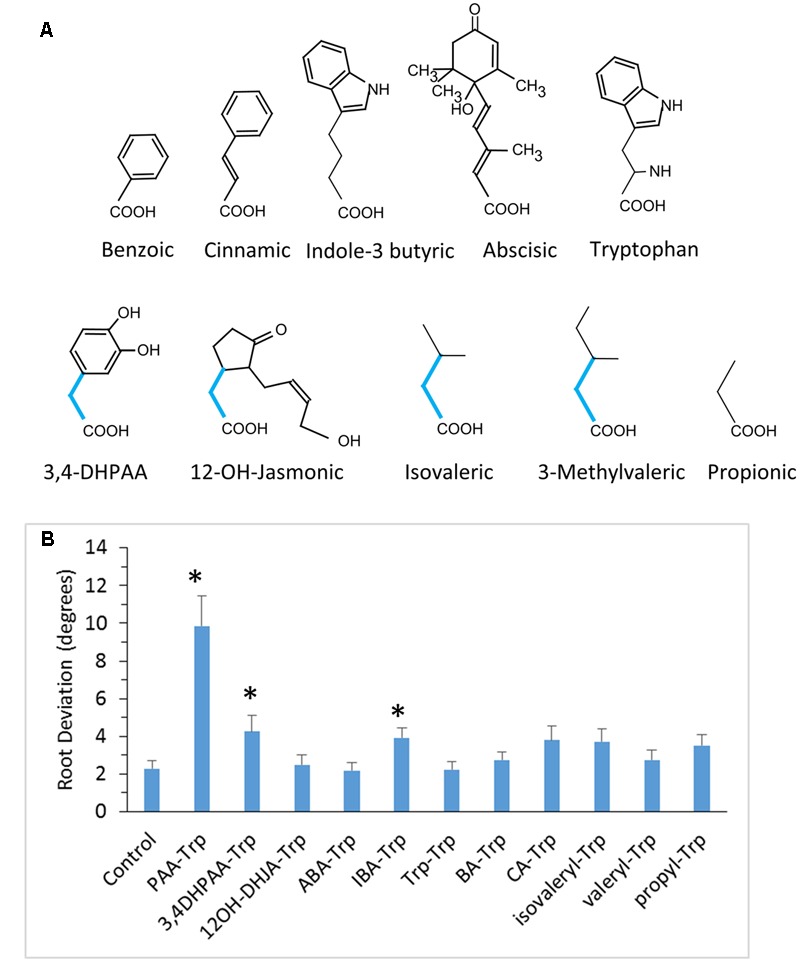
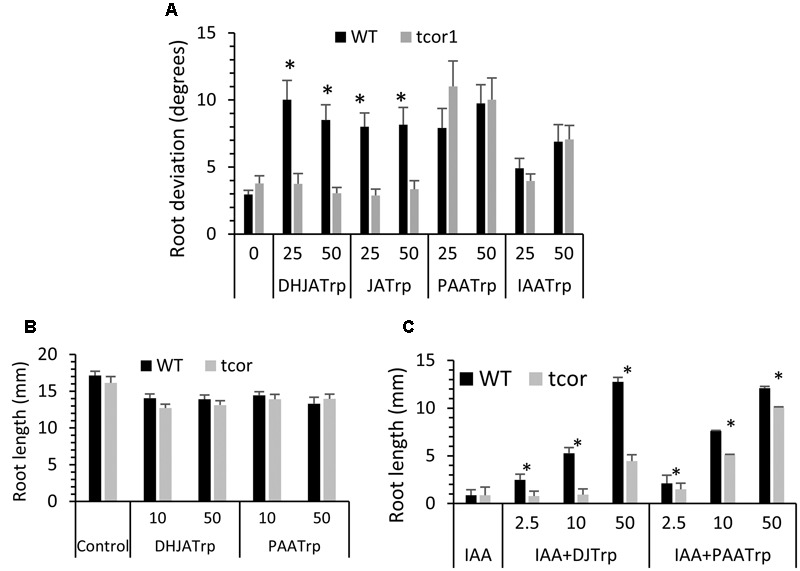
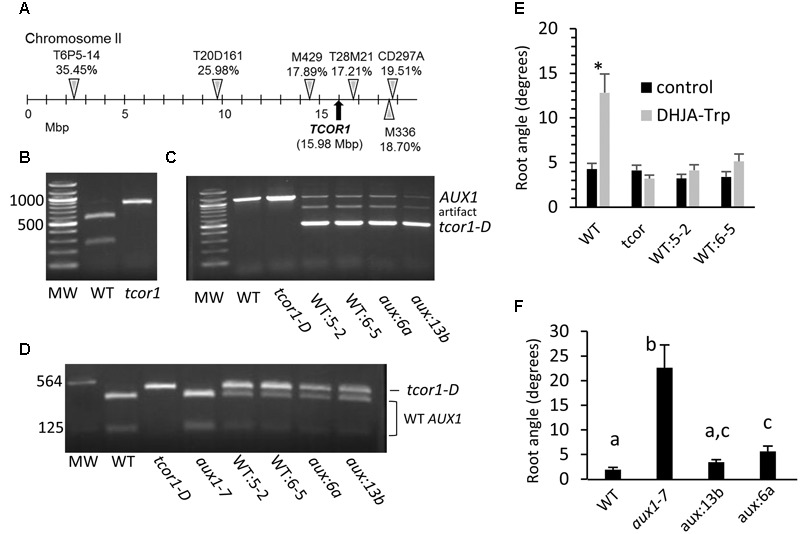
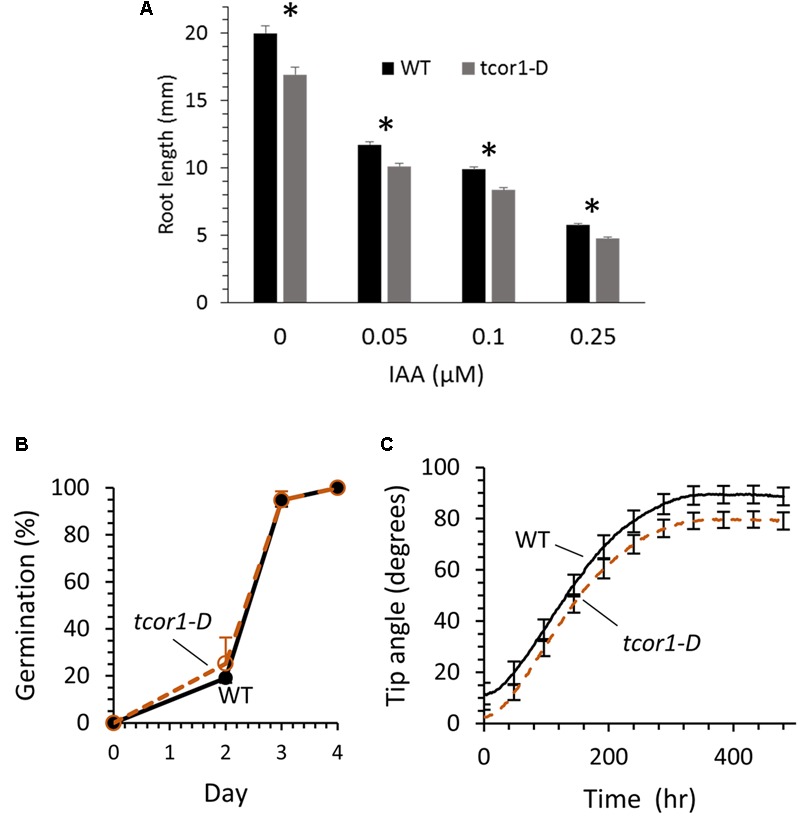
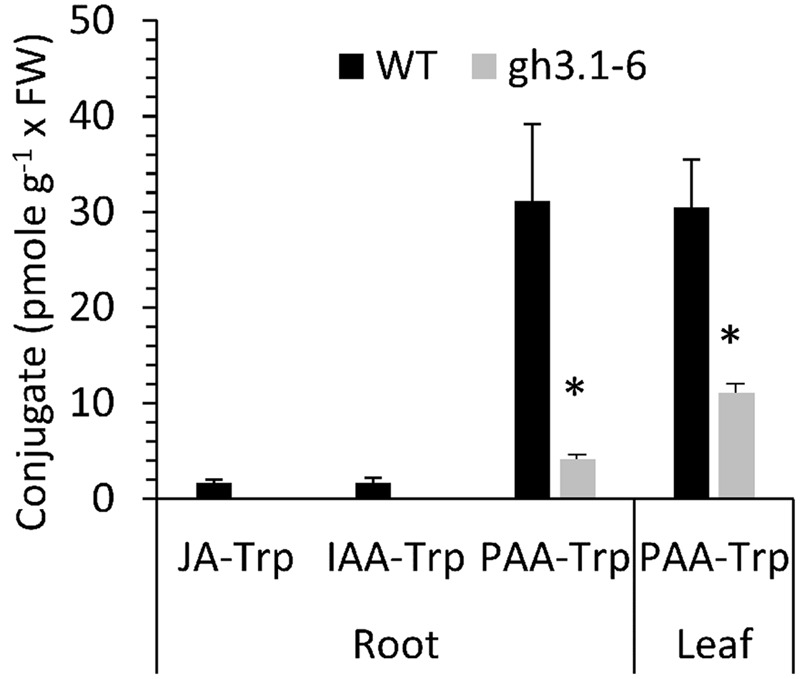
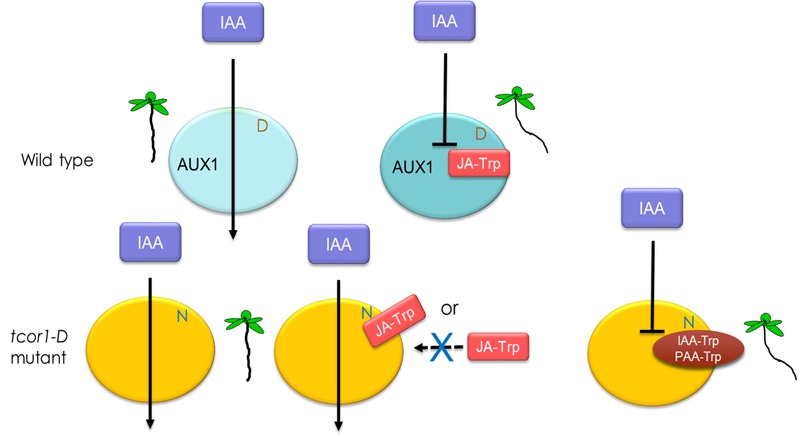
Amide-linked conjugates between tryptophan (Trp) and jasmonic (JA) or indole-3-acetic (IAA) acids interfered with gravitropism and other auxin-dependent activities in Arabidopsis, but the mechanism was unclear. To identify structural features necessary for activity several additional Trp conjugates were synthesized. The phenylacetic acid (PAA) conjugate was active, while several others were not. Common features of active conjugates is that they have ring structures that are linked to Trp through an acetic acid side chain, while longer or shorter linkages are inactive or less active. A dominant mutant, called tryptophan conjugate response1-D that is insensitive to JA-Trp, but still sensitive to other active conjugates, was identified and the defect was found to be a substitution of Asn for Asp456 in the C-terminal domain of the IAA cellular permease AUX1. Mutant seedling primary root growth in the absence of added conjugate was 15% less than WT, but otherwise plant phenotype appeared normal. These results suggest that JA-Trp disrupts AUX1 activity, but that endogenous JA-Trp has only a minor role in regulating plant growth. In contrast with IAA- and JA-Trp, which are present at <2 pmole g-1 FW, PAA-Trp was found at about 30 pmole g-1 FW. The latter, or other undiscovered Trp conjugates, may still have important endogenous roles, possibly helping to coordinate other pathways with auxin response.
Keywords: AUX1; auxin; conjugate; gravitropism; indole-3-acetic acid; jasmonic acid; phenylacetic acid; tryptophan.
Figures

References
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases