

Mechanism of bacterial gene rearrangement: SprA-catalyzed precise DNA recombination and its directionality control by SprB ensure the gene rearrangement and stable expression of spsM during sporulation in Bacillus subtilis
- PMID: 28535266
- PMCID: PMC5499854
- DOI: 10.1093/nar/gkx466
Mechanism of bacterial gene rearrangement: SprA-catalyzed precise DNA recombination and its directionality control by SprB ensure the gene rearrangement and stable expression of spsM during sporulation in Bacillus subtilis
Abstract
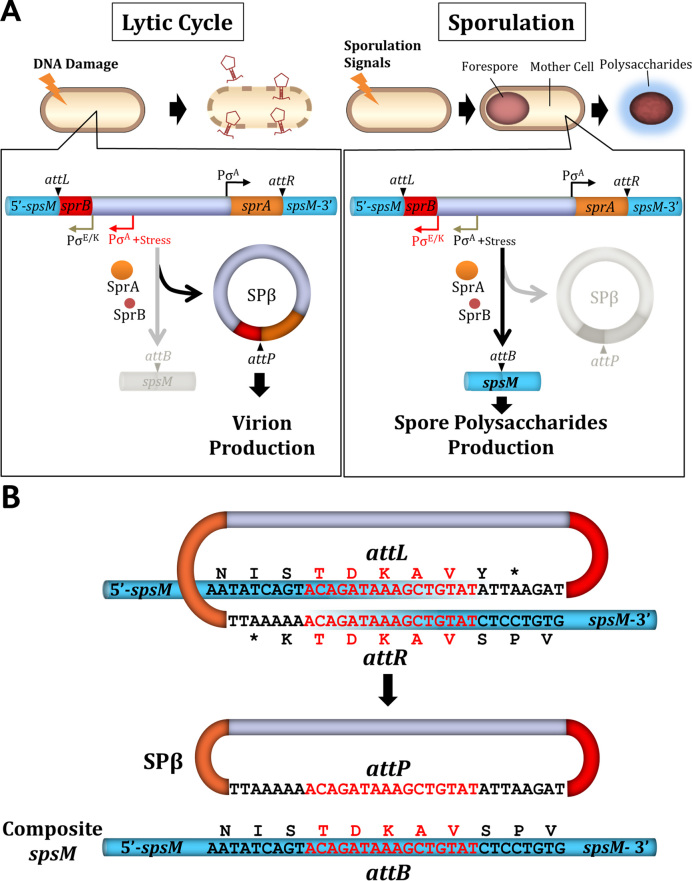
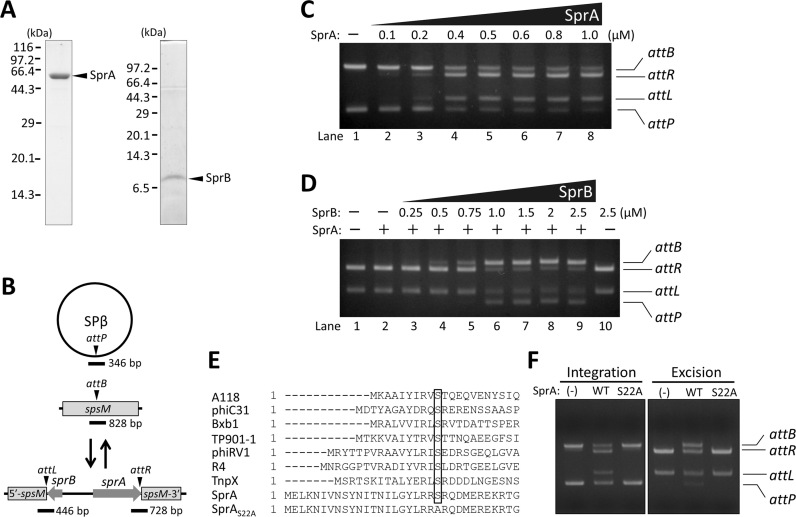
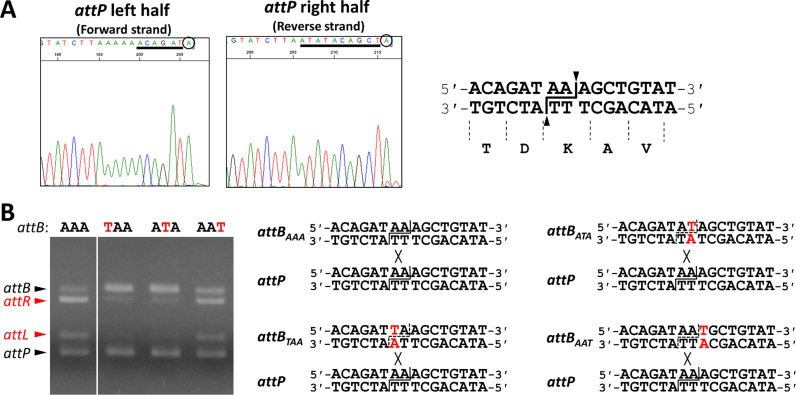
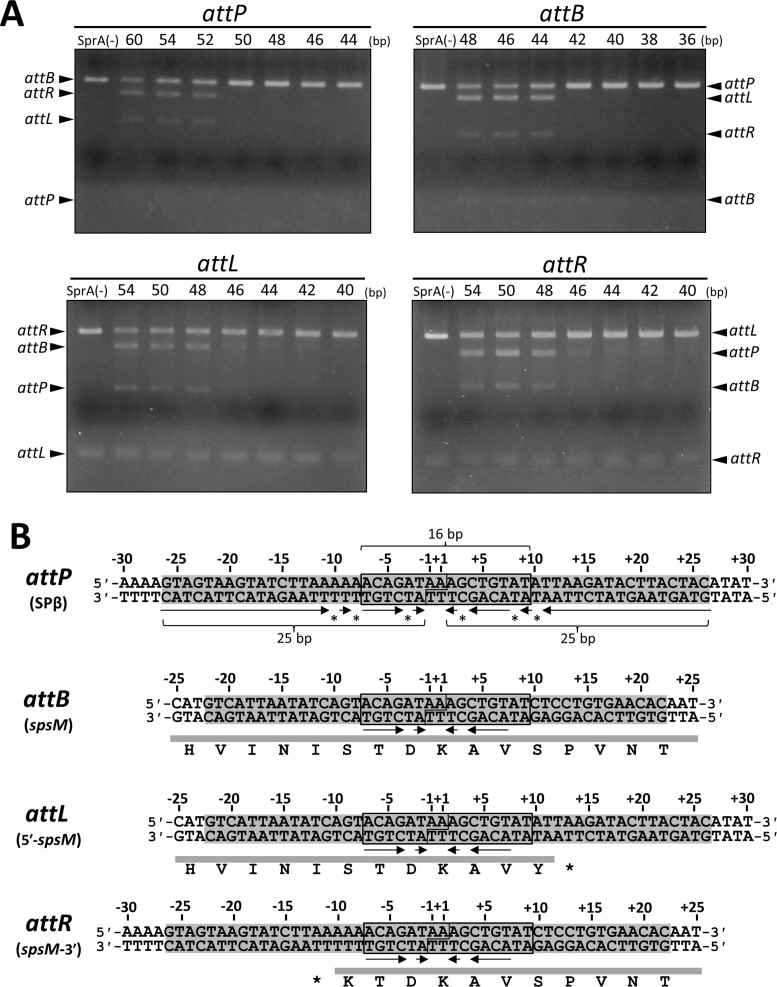
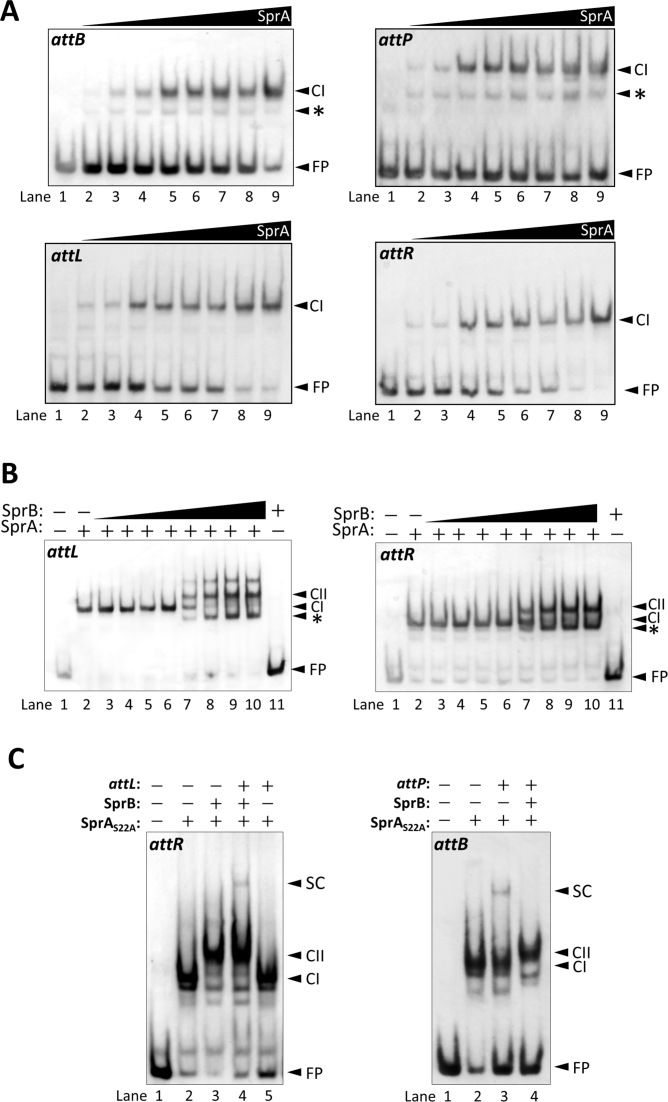
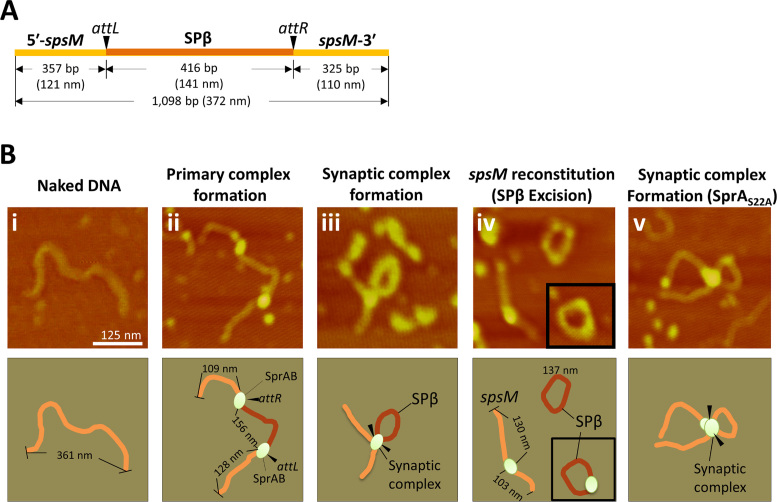
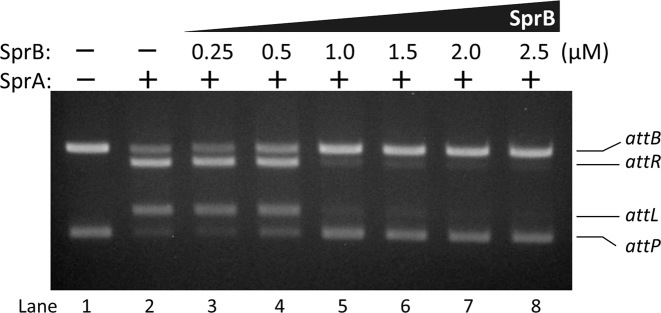
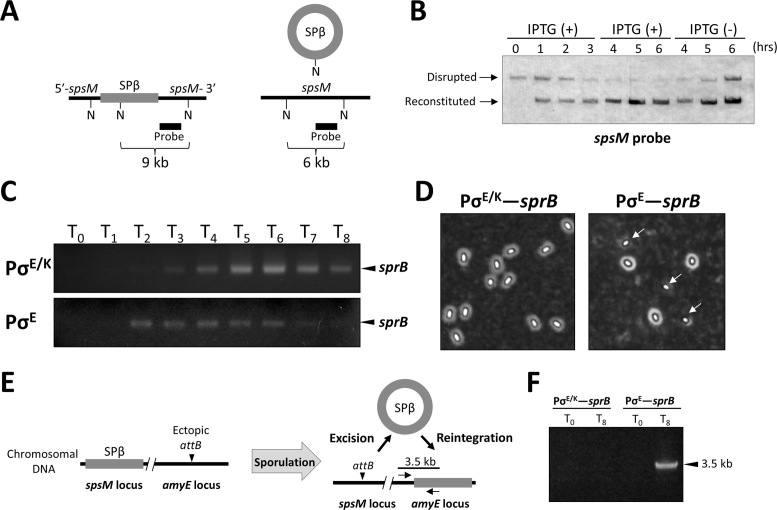
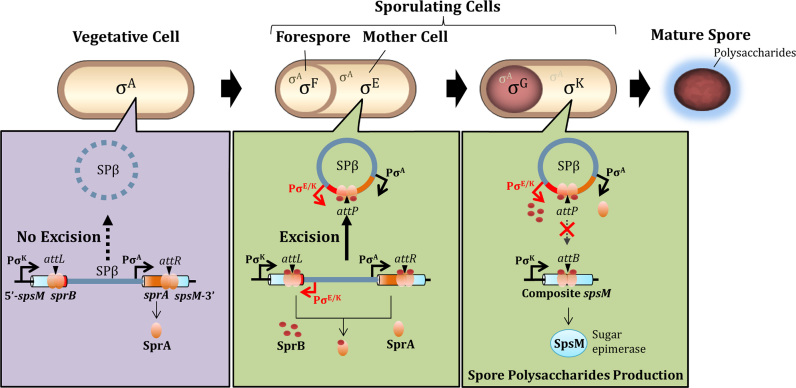
A sporulation-specific gene, spsM, is disrupted by an active prophage, SPβ, in the genome of Bacillus subtilis. SPβ excision is required for two critical steps: the onset of the phage lytic cycle and the reconstitution of the spsM-coding frame during sporulation. Our in vitro study demonstrated that SprA, a serine-type integrase, catalyzed integration and excision reactions between attP of SPβ and attB within spsM, while SprB, a recombination directionality factor, was necessary only for the excision between attL and attR in the SPβ lysogenic chromosome. DNA recombination occurred at the center of the short inverted repeat motif in the unique conserved 16 bp sequence among the att sites (5΄-ACAGATAA/AGCTGTAT-3΄; slash, breakpoint; underlines, inverted repeat), where SprA produced the 3΄-overhanging AA and TT dinucleotides for rejoining the DNA ends through base-pairing. Electrophoretic mobility shift assay showed that SprB promoted synapsis of SprA subunits bound to the two target sites during excision but impaired it during integration. In vivo data demonstrated that sprB expression that lasts until the late stage of sporulation is crucial for stable expression of reconstituted spsM without reintegration of the SPβ prophage. These results present a deeper understanding of the mechanism of the prophage-mediated bacterial gene regulatory system.
© The Author(s) 2017. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures
References
-
- Sakano H., Rogers J.H., Huppi K., Brack C., Traunecker A., Maki R., Wall R., Tonegawa S.. Domains and the hinge region of an immunoglobulin heavy chain are encoded in separate DNA segments. Nature. 1979; 277:627–633. - PubMed
-
- Schatz D.G., Ji Y.. Recombination centres and the orchestration of V(D)J recombination. Nat. Rev. Immunol. 2011; 11:251–263. - PubMed
-
- Schatz D.G., Oettinger M.A., Baltimore D.. The V(D)J recombination activating gene, RAG-1. Cell. 1989; 59:1035–1048. - PubMed
-
- Oettinger M.A., Schatz D.G., Gorka C., Baltimore D.. RAG-1 and RAG-2, adjacent genes that synergistically activate V(D)J recombination. Science. 1990; 248:1517–1523. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
