

Dephosphorylation of the Ndc80 Tail Stabilizes Kinetochore-Microtubule Attachments via the Ska Complex
- PMID: 28535376
- PMCID: PMC5572820
- DOI: 10.1016/j.devcel.2017.04.013
Dephosphorylation of the Ndc80 Tail Stabilizes Kinetochore-Microtubule Attachments via the Ska Complex
Abstract
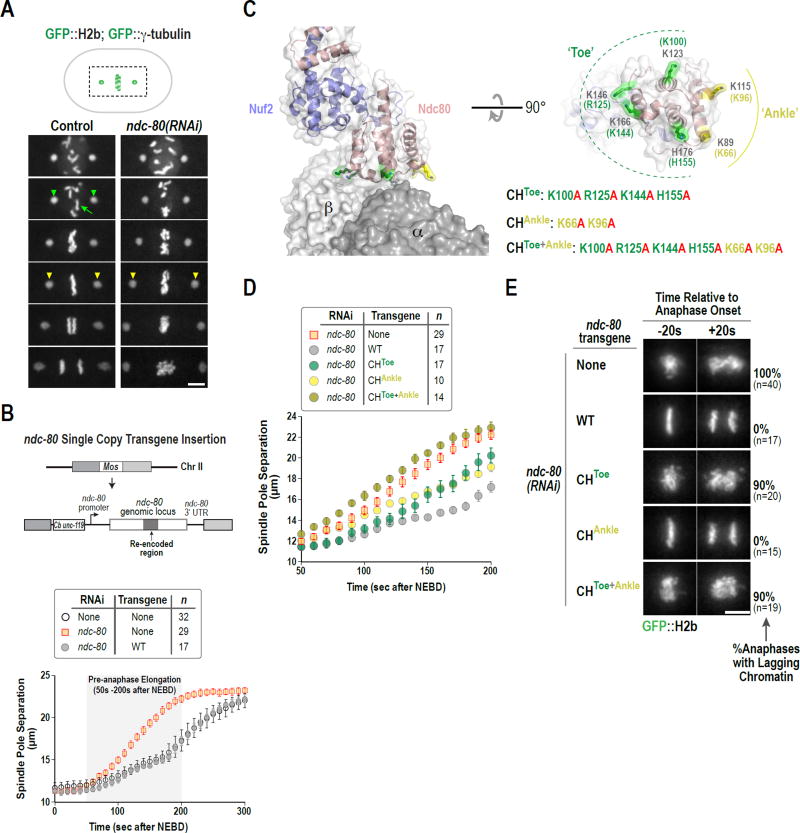
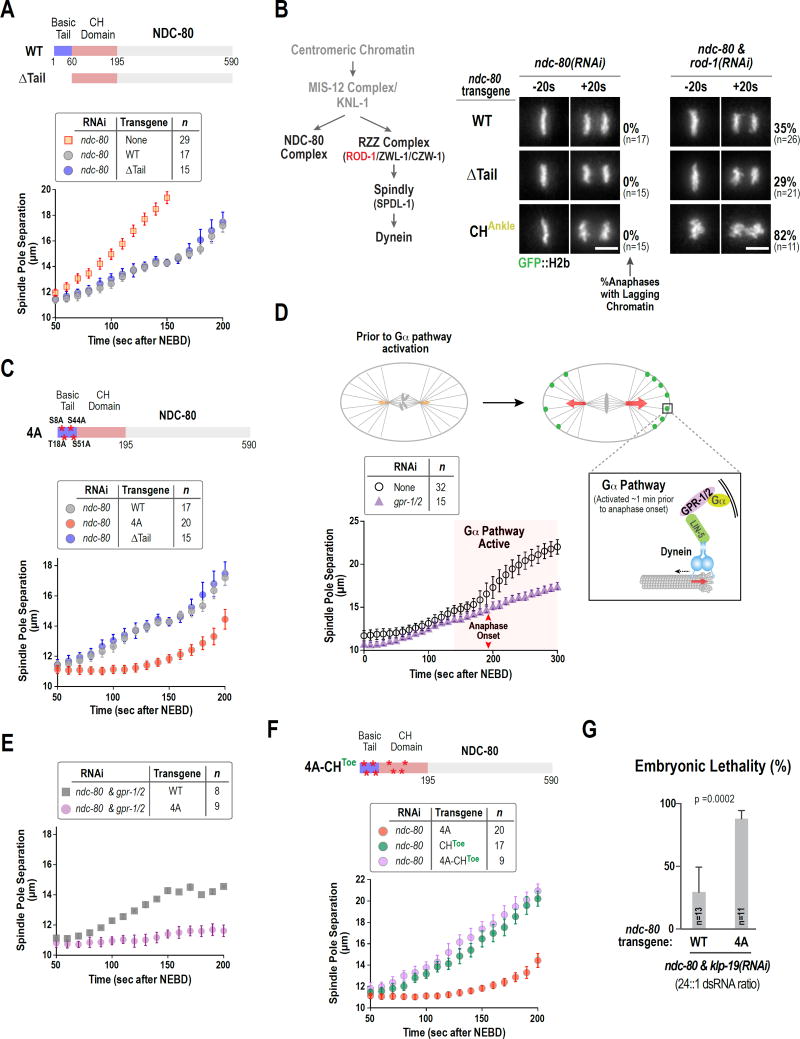
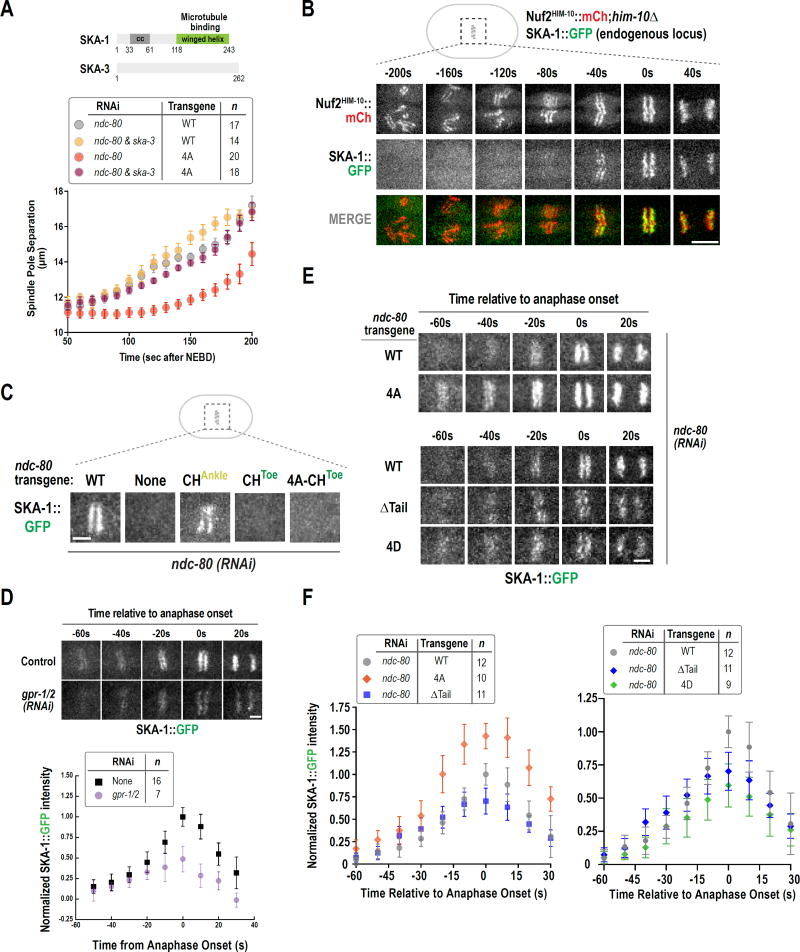
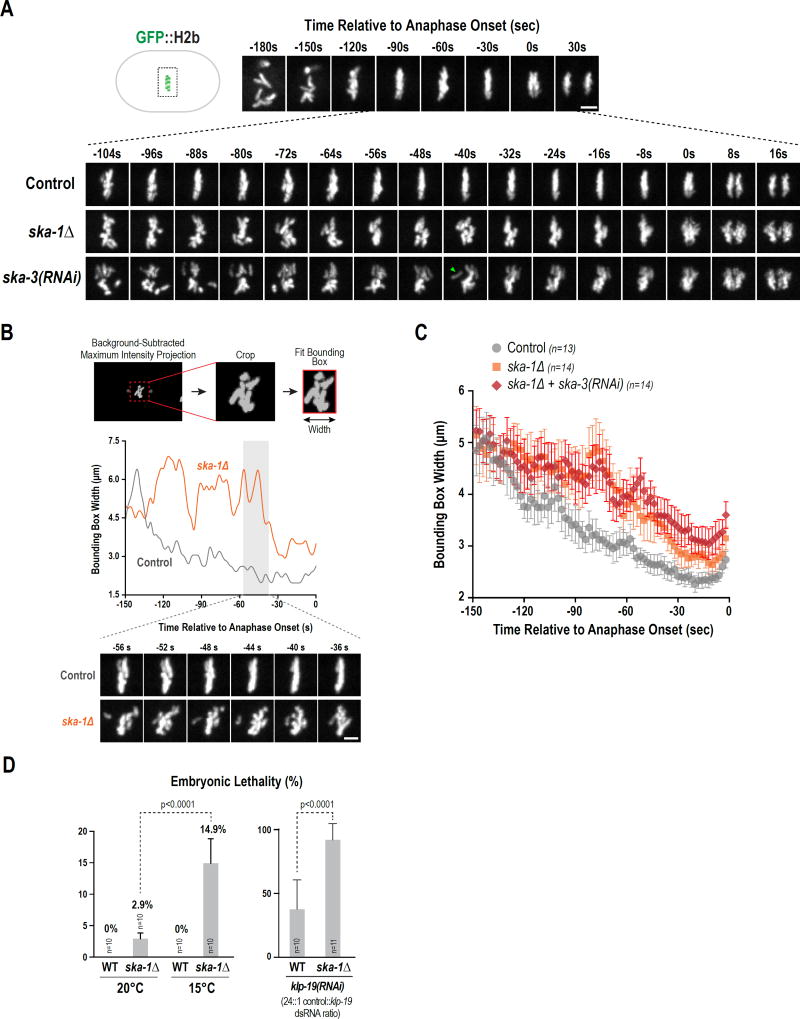
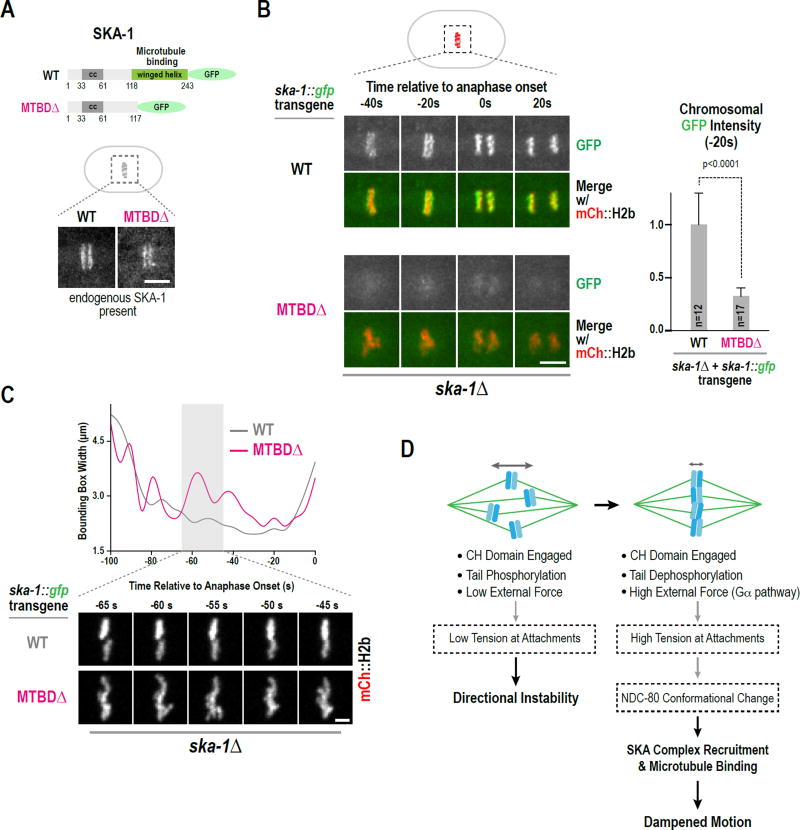
During cell division, genome inheritance is orchestrated by microtubule attachments formed at kinetochores of mitotic chromosomes. The primary microtubule coupler at the kinetochore, the Ndc80 complex, is regulated by Aurora kinase phosphorylation of its N-terminal tail. Dephosphorylation is proposed to stabilize kinetochore-microtubule attachments by strengthening electrostatic interactions of the tail with the microtubule lattice. Here, we show that removal of the Ndc80 tail, which compromises in vitro microtubule binding, has no effect on kinetochore-microtubule attachments in the Caenorhabditis elegans embryo. Despite this, preventing Aurora phosphorylation of the tail results in prematurely stable attachments that restrain spindle elongation. This premature stabilization requires the conserved microtubule-binding Ska complex, which enriches at attachment sites prior to anaphase onset to dampen chromosome motion. We propose that Ndc80-tail dephosphorylation promotes stabilization of kinetochore-microtubule attachments via the Ska complex and that this mechanism ensures accurate segregation by constraining chromosome motion following biorientation on the spindle.
Keywords: Ndc80 complex; Ska complex; cell division; cell polarity; centromere; chromosome segregation; kinetochore; microtubule; mitosis.
Copyright © 2017 Elsevier Inc. All rights reserved.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
