

Structural Basis for Asymmetric Conductance of the Influenza M2 Proton Channel Investigated by Solid-State NMR Spectroscopy
- PMID: 28535993
- PMCID: PMC5761075
- DOI: 10.1016/j.jmb.2017.05.015
Structural Basis for Asymmetric Conductance of the Influenza M2 Proton Channel Investigated by Solid-State NMR Spectroscopy
Abstract
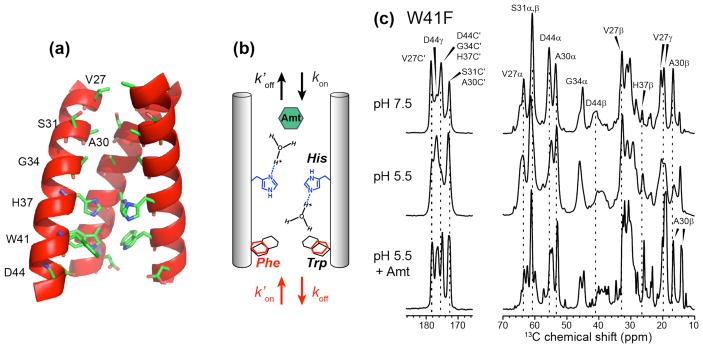
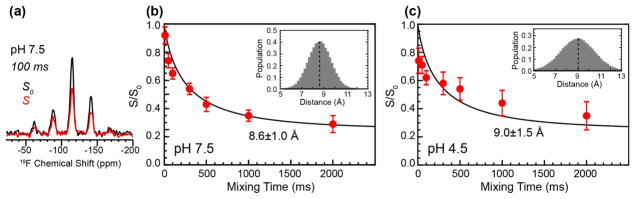
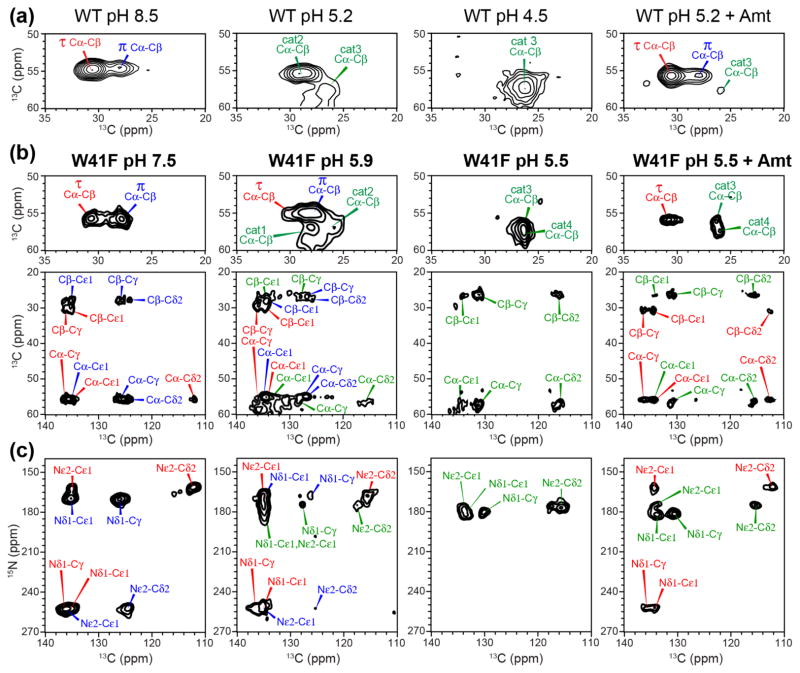
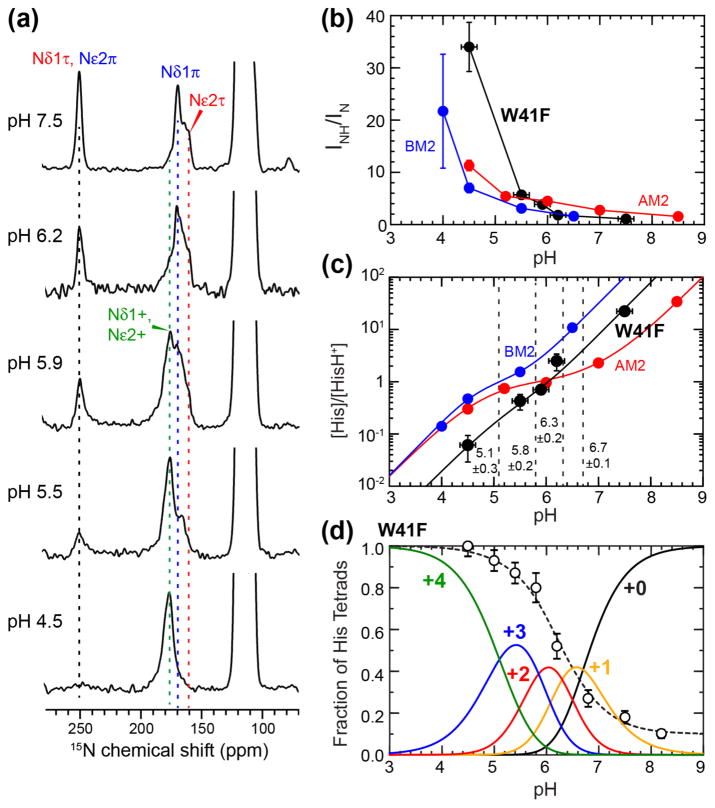
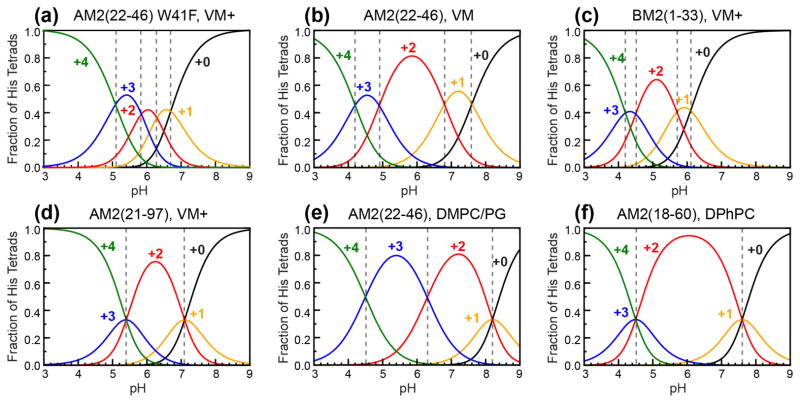
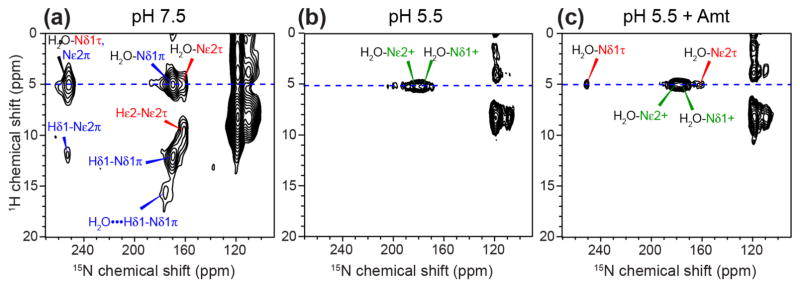
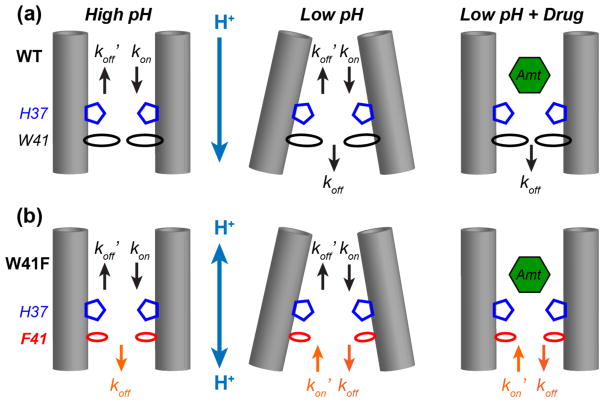
The influenza M2 protein forms an acid-activated proton channel that is essential for virus replication. The transmembrane H37 selects for protons under low external pH while W41 ensures proton conduction only from the N terminus to the C terminus and prevents reverse current under low internal pH. Here, we address the molecular basis for this asymmetric conduction by investigating the structure and dynamics of a mutant channel, W41F, which permits reverse current under low internal pH. Solid-state NMR experiments show that W41F M2 retains the pH-dependent α-helical conformations and tetrameric structure of the wild-type (WT) channel but has significantly altered protonation and tautomeric equilibria at H37. At high pH, the H37 structure is shifted toward the π tautomer and less cationic tetrads, consistent with faster forward deprotonation to the C terminus. At low pH, the mutant channel contains more cationic tetrads than the WT channel, consistent with faster reverse protonation from the C terminus. 15N NMR spectra allow the extraction of four H37 pKas and show that the pKas are more clustered in the mutant channel compared to WT M2. Moreover, binding of the antiviral drug, amantadine, at the N-terminal pore at low pH did not convert all histidines to the neutral state, as seen in WT M2, but left half of all histidines cationic, unambiguously demonstrating C-terminal protonation of H37 in the mutant. These results indicate that asymmetric conduction in WT M2 is due to W41 inhibition of C-terminal acid activation by H37. When Trp is replaced by Phe, protons can be transferred to H37 bidirectionally with distinct rate constants.
Keywords: gating; ion channels; magic-angle-spinning NMR; proton dissociation equilibrium; tautomeric equilibrium.
Copyright © 2017 Elsevier Ltd. All rights reserved.
Figures


References
-
- Hille B. Ionic channels of excitable membranes. 2. Sundeland: Sinauer Associates Inc; 1992.
-
- Pinto LH, Lamb RA. The M2 proton channels of influenza A and B viruses. J Biol Chem. 2006;281:8997–9000. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
