

The LOTUS domain is a conserved DEAD-box RNA helicase regulator essential for the recruitment of Vasa to the germ plasm and nuage
- PMID: 28536148
- PMCID: PMC5458760
- DOI: 10.1101/gad.297051.117
The LOTUS domain is a conserved DEAD-box RNA helicase regulator essential for the recruitment of Vasa to the germ plasm and nuage
Abstract
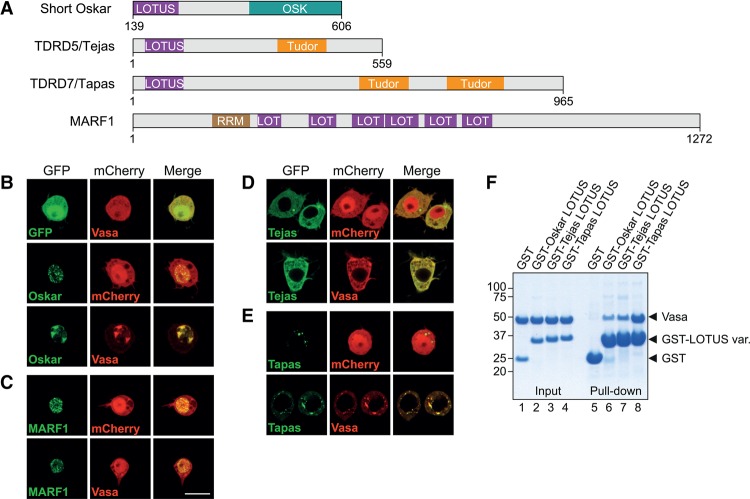
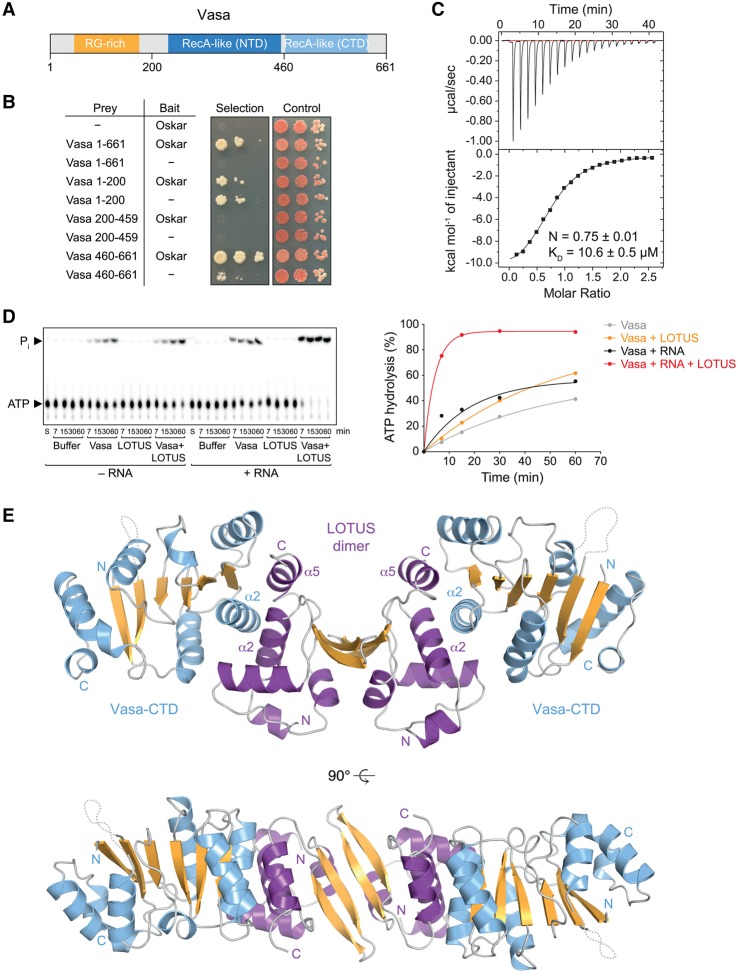
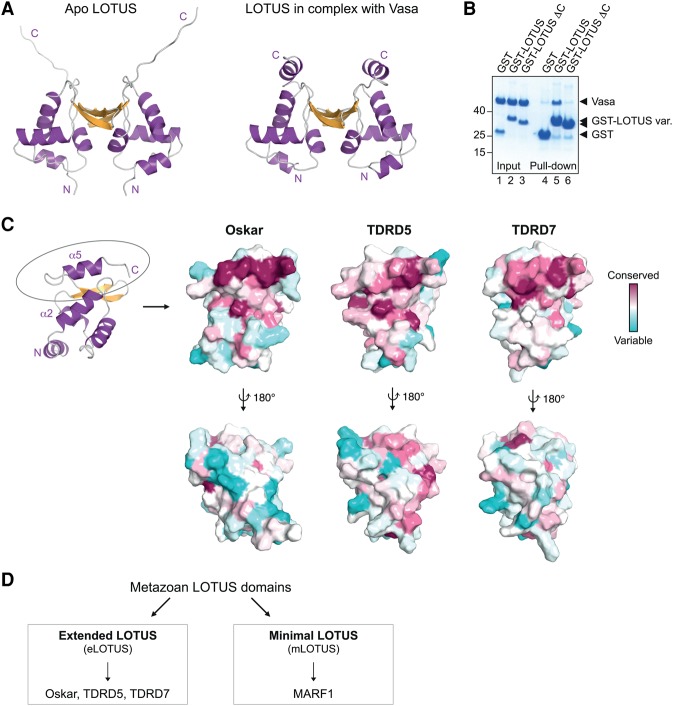
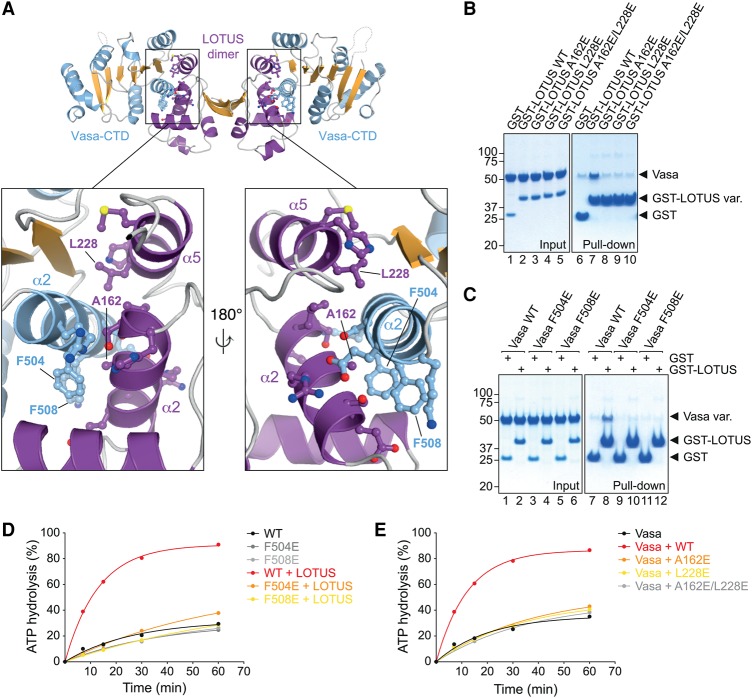
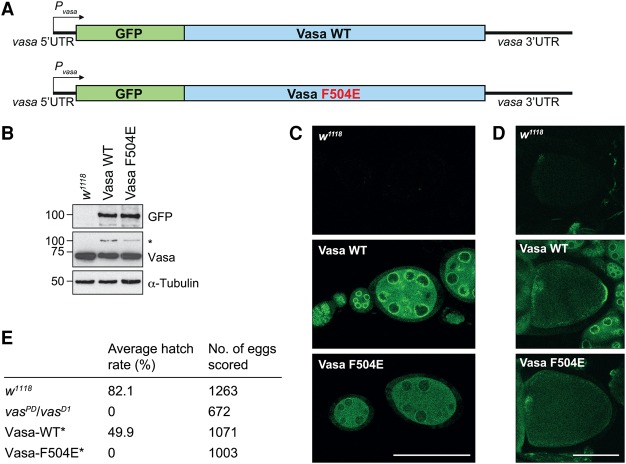
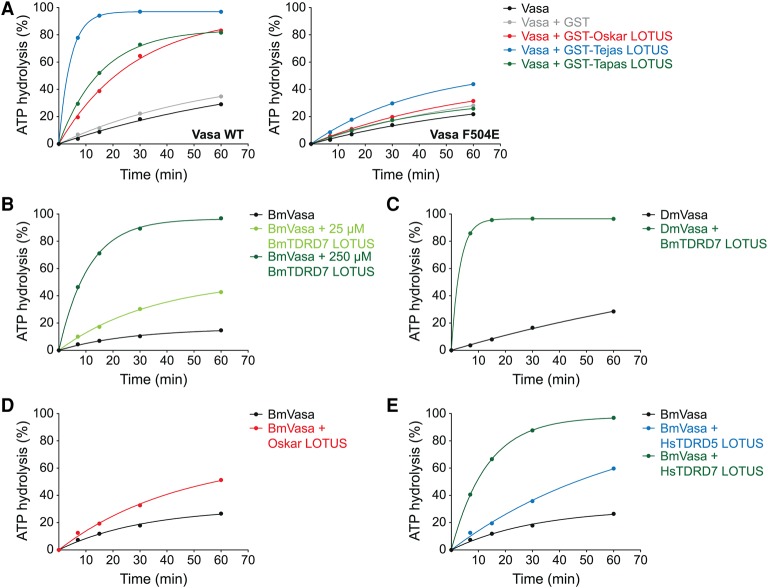
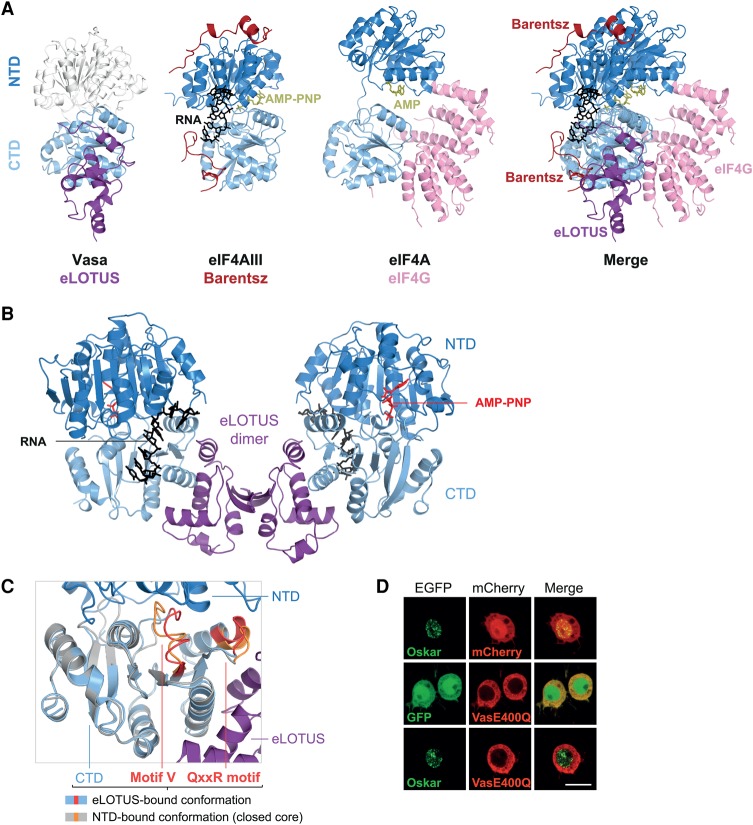
DEAD-box RNA helicases play important roles in a wide range of metabolic processes. Regulatory proteins can stimulate or block the activity of DEAD-box helicases. Here, we show that LOTUS (Limkain, Oskar, and Tudor containing proteins 5 and 7) domains present in the germline proteins Oskar, TDRD5 (Tudor domain-containing 5), and TDRD7 bind and stimulate the germline-specific DEAD-box RNA helicase Vasa. Our crystal structure of the LOTUS domain of Oskar in complex with the C-terminal RecA-like domain of Vasa reveals that the LOTUS domain occupies a surface on a DEAD-box helicase not implicated previously in the regulation of the enzyme's activity. We show that, in vivo, the localization of Drosophila Vasa to the nuage and germ plasm depends on its interaction with LOTUS domain proteins. The binding and stimulation of Vasa DEAD-box helicases by LOTUS domains are widely conserved.
Keywords: DEAD-box RNA helicase Vasa; Oskar; TDRD5; TDRD7; germ plasm; nuage.
© 2017 Jeske et al.; Published by Cold Spring Harbor Laboratory Press.
Figures
Similar articles
-
The Crystal Structure of the Drosophila Germline Inducer Oskar Identifies Two Domains with Distinct Vasa Helicase- and RNA-Binding Activities.Cell Rep. 2015 Jul 28;12(4):587-98. doi: 10.1016/j.celrep.2015.06.055. Epub 2015 Jul 16. Cell Rep. 2015. PMID: 26190108
-
Germ plasm localisation of the HELICc of Vasa in Drosophila: analysis of domain sufficiency and amino acids critical for localisation.Sci Rep. 2015 Sep 30;5:14703. doi: 10.1038/srep14703. Sci Rep. 2015. PMID: 26419889 Free PMC article.
-
Targeting and anchoring Tudor in the pole plasm of the Drosophila oocyte.PLoS One. 2010 Dec 15;5(12):e14362. doi: 10.1371/journal.pone.0014362. PLoS One. 2010. PMID: 21179512 Free PMC article.
-
The DEAD-box helicase Vasa: evidence for a multiplicity of functions in RNA processes and developmental biology.Biochim Biophys Acta. 2013 Aug;1829(8):810-6. doi: 10.1016/j.bbagrm.2013.04.005. Epub 2013 Apr 13. Biochim Biophys Acta. 2013. PMID: 23587717 Review.
-
LOTUS-domain proteins - developmental effectors from a molecular perspective.Biol Chem. 2020 Nov 4;402(1):7-23. doi: 10.1515/hsz-2020-0270. Print 2020 Nov 18. Biol Chem. 2020. PMID: 33544496 Review.
Cited by
-
Patterning the Drosophila embryo: A paradigm for RNA-based developmental genetic regulation.Wiley Interdiscip Rev RNA. 2020 Nov;11(6):e1610. doi: 10.1002/wrna.1610. Epub 2020 Jun 15. Wiley Interdiscip Rev RNA. 2020. PMID: 32543002 Free PMC article. Review.
-
Highly Overexpressed AtC3H18 Impairs Microgametogenesis via Promoting the Continuous Assembly of mRNP Granules.Front Plant Sci. 2022 Jul 15;13:932793. doi: 10.3389/fpls.2022.932793. eCollection 2022. Front Plant Sci. 2022. PMID: 35909782 Free PMC article.
-
The Caenorhabditis elegans TDRD5/7-like protein, LOTR-1, interacts with the helicase ZNFX-1 to balance epigenetic signals in the germline.PLoS Genet. 2022 Jun 3;18(6):e1010245. doi: 10.1371/journal.pgen.1010245. eCollection 2022 Jun. PLoS Genet. 2022. PMID: 35657999 Free PMC article.
-
Bucky Ball Is a Novel Zebrafish Vasa ATPase Activator.Biomolecules. 2021 Oct 13;11(10):1507. doi: 10.3390/biom11101507. Biomolecules. 2021. PMID: 34680140 Free PMC article.
-
Proteins rather than mRNAs regulate nucleation and persistence of Oskar germ granules in Drosophila.Cell Rep. 2023 Jul 25;42(7):112723. doi: 10.1016/j.celrep.2023.112723. Epub 2023 Jun 28. Cell Rep. 2023. PMID: 37384531 Free PMC article.
References
-
- Andersen CBF, Ballut L, Johansen JS, Chamieh H, Nielsen KH, Oliveira CLP, Pedersen JS, Séraphin B, Le Hir H, Andersen GR. 2006. Structure of the exon junction core complex with a trapped DEAD-box ATPase bound to RNA. Science 313: 1968–1972. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials