

Exosomes from uninfected cells activate transcription of latent HIV-1
- PMID: 28536264
- PMCID: PMC5512065
- DOI: 10.1074/jbc.M117.793521
Exosomes from uninfected cells activate transcription of latent HIV-1
Erratum in
-
Exosomes from uninfected cells activate transcription of latent HIV-1.J Biol Chem. 2017 Sep 8;292(36):14764. doi: 10.1074/jbc.A117.793521. J Biol Chem. 2017. PMID: 28887434 Free PMC article. No abstract available.
Abstract
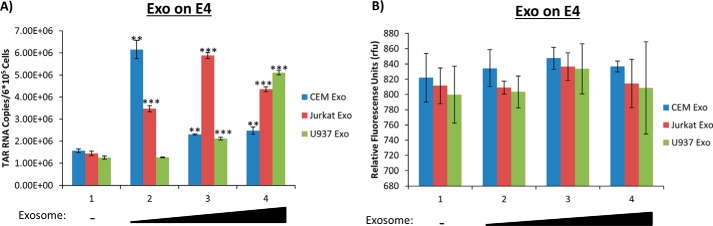
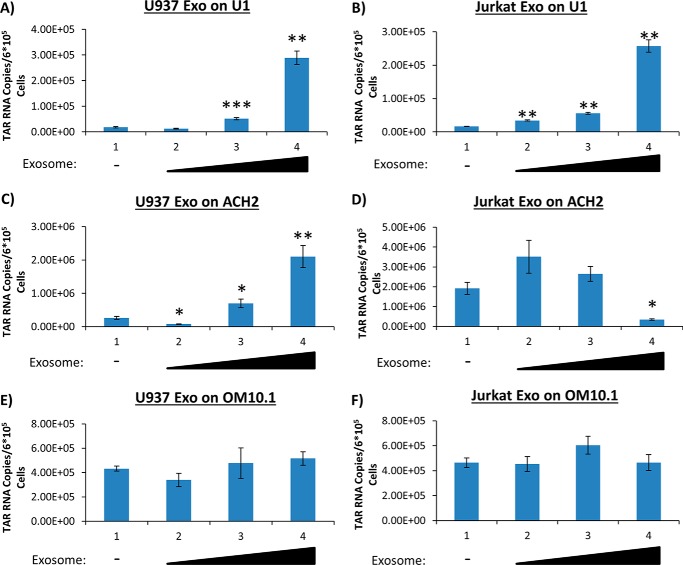
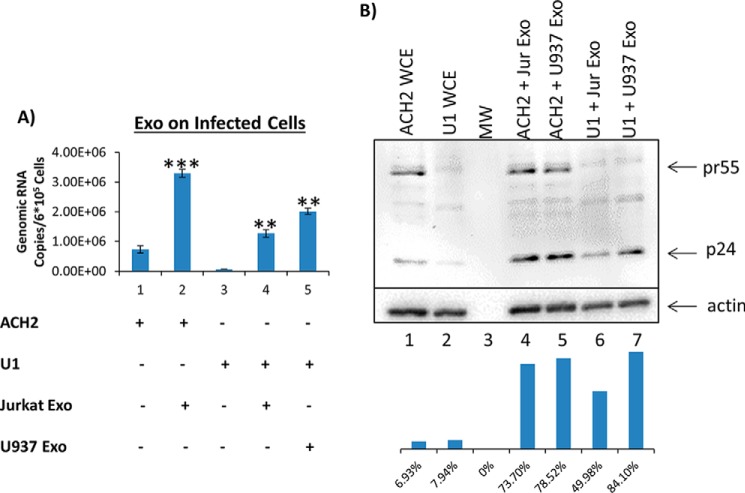
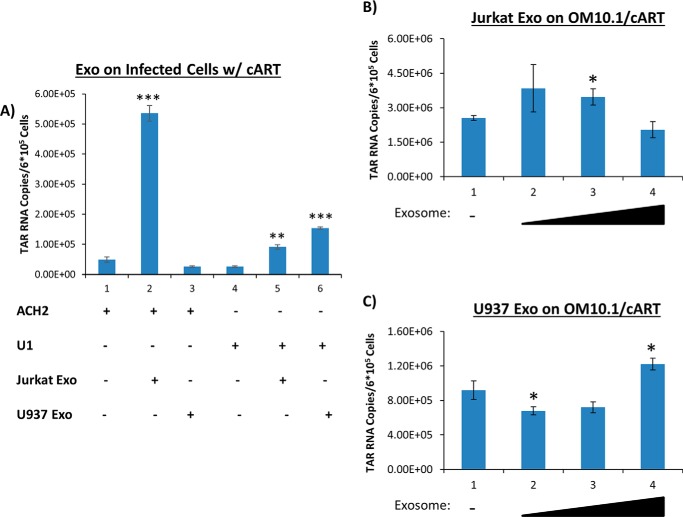
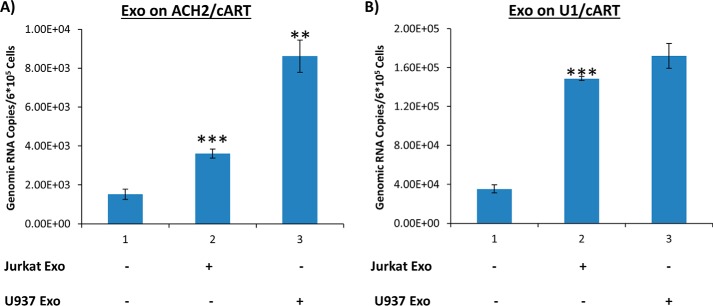
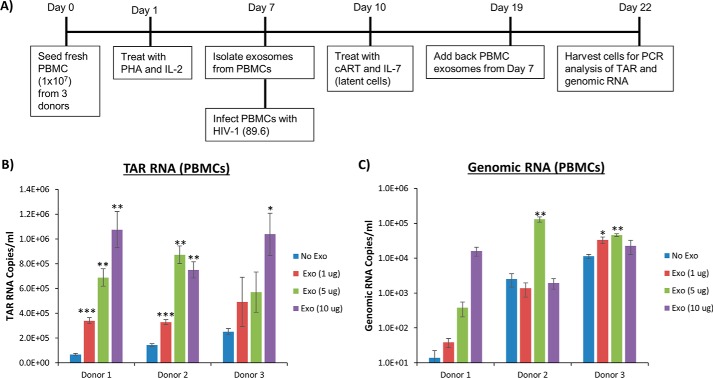
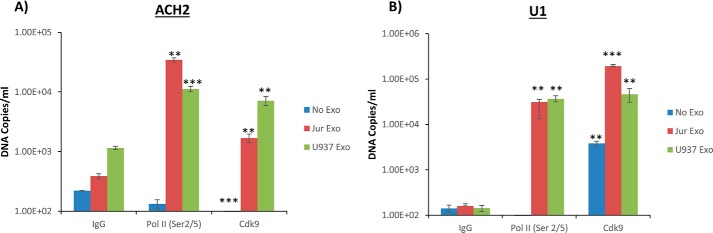
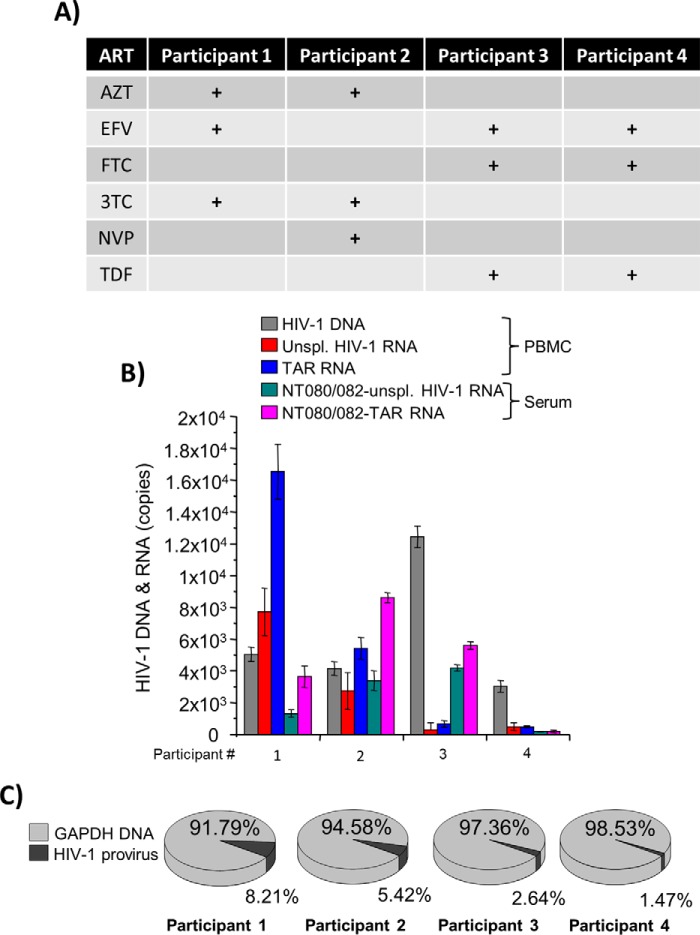
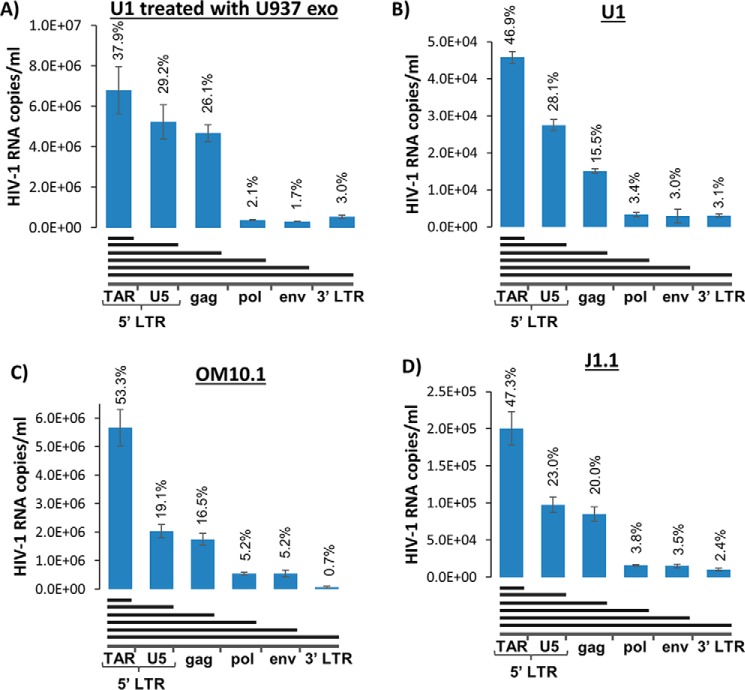
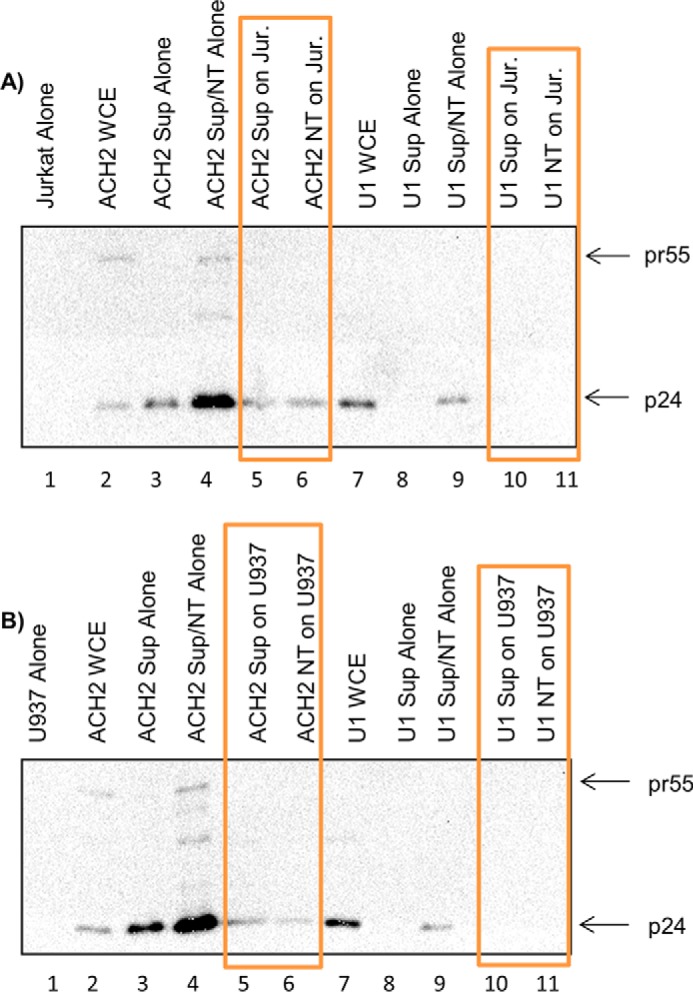
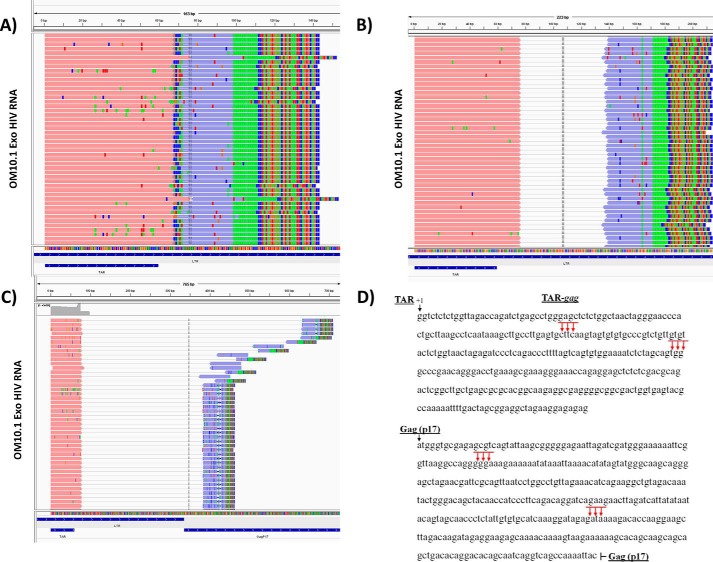
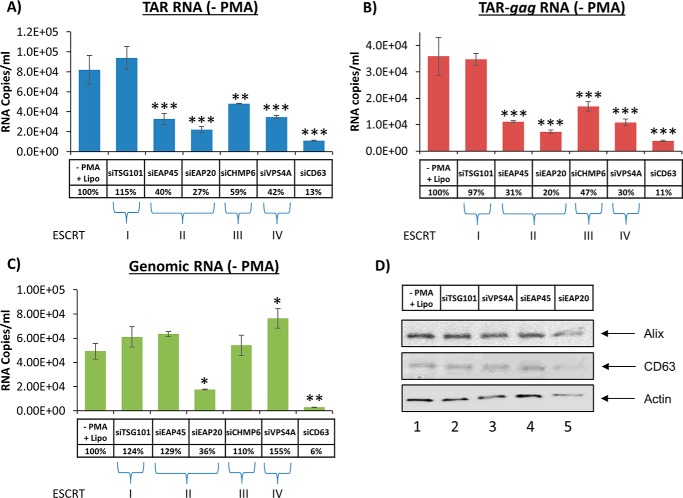
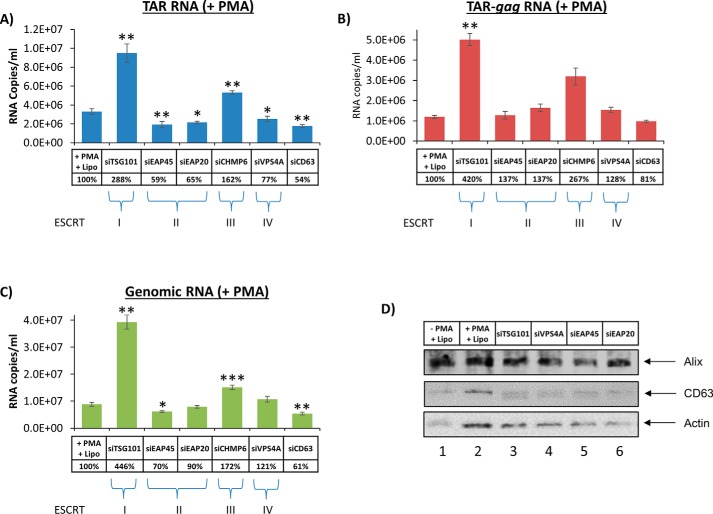
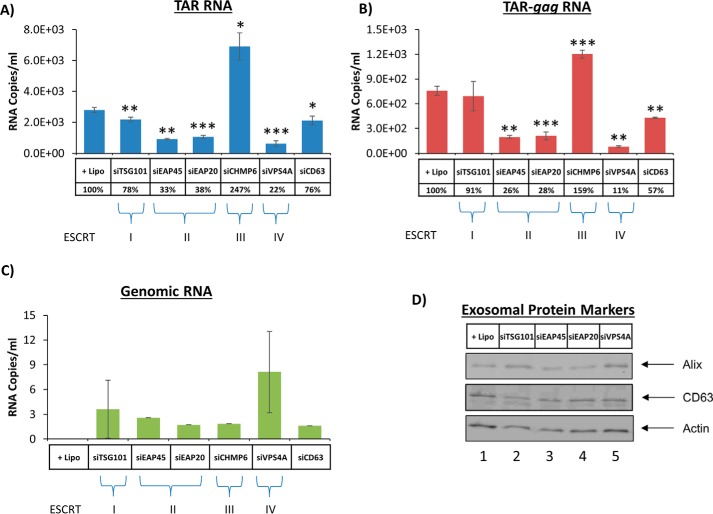
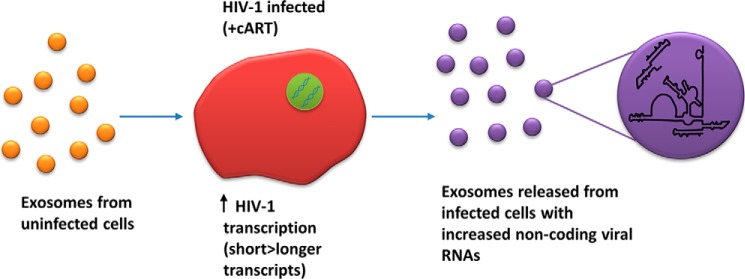
HIV-1 infection causes AIDS, infecting millions worldwide. The virus can persist in a state of chronic infection due to its ability to become latent. We have previously shown a link between HIV-1 infection and exosome production. Specifically, we have reported that exosomes transport viral proteins and RNA from infected cells to neighboring uninfected cells. These viral products could then elicit an innate immune response, leading to activation of the Toll-like receptor and NF-κB pathways. In this study, we asked whether exosomes from uninfected cells could activate latent HIV-1 in infected cells. We observed that irrespective of combination antiretroviral therapy, both short- and long-length viral transcripts were increased in wild-type HIV-1-infected cells exposed to purified exosomes from uninfected cells. A search for a possible mechanism for this finding revealed that the exosomes increase RNA polymerase II loading onto the HIV-1 promoter in the infected cells. These viral transcripts, which include trans-activation response (TAR) RNA and a novel RNA that we termed TAR-gag, can then be packaged into exosomes and potentially be exported to neighboring uninfected cells, leading to increased cellular activation. To better decipher the exosome release pathways involved, we used siRNA to suppress expression of ESCRT (endosomal sorting complex required for transport) proteins and found that ESCRT II and IV significantly control exosome release. Collectively, these results imply that exosomes from uninfected cells activate latent HIV-1 in infected cells and that true transcriptional latency may not be possible in vivo, especially in the presence of combination antiretroviral therapy.
Keywords: T-cell; endosomal sorting complexes required for transport (ESCRT); exosome (vesicle); human immunodeficiency virus (HIV); latency; monocyte; small interfering RNA (siRNA); transcription.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
