

Modulation of Aneuploidy in Leishmania donovani during Adaptation to Different In Vitro and In Vivo Environments and Its Impact on Gene Expression
- PMID: 28536289
- PMCID: PMC5442457
- DOI: 10.1128/mBio.00599-17
Modulation of Aneuploidy in Leishmania donovani during Adaptation to Different In Vitro and In Vivo Environments and Its Impact on Gene Expression
Abstract
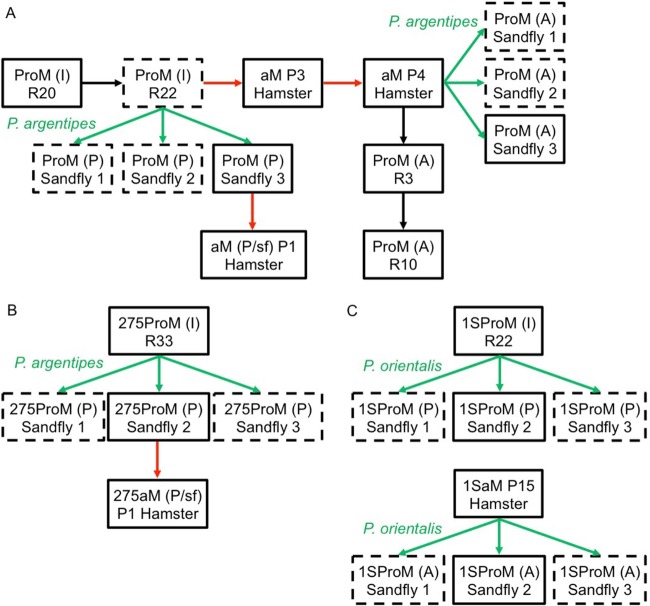
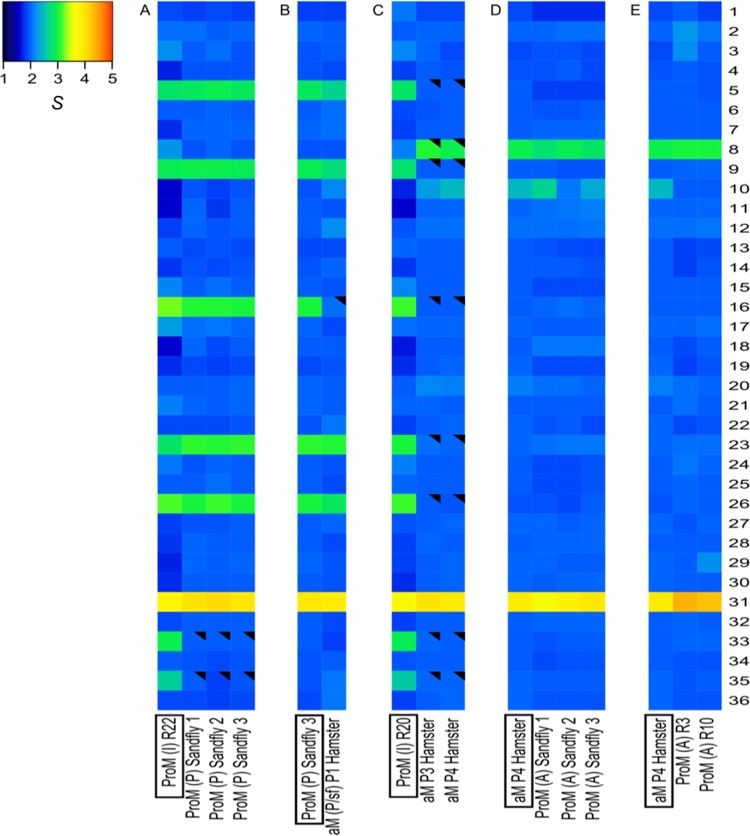
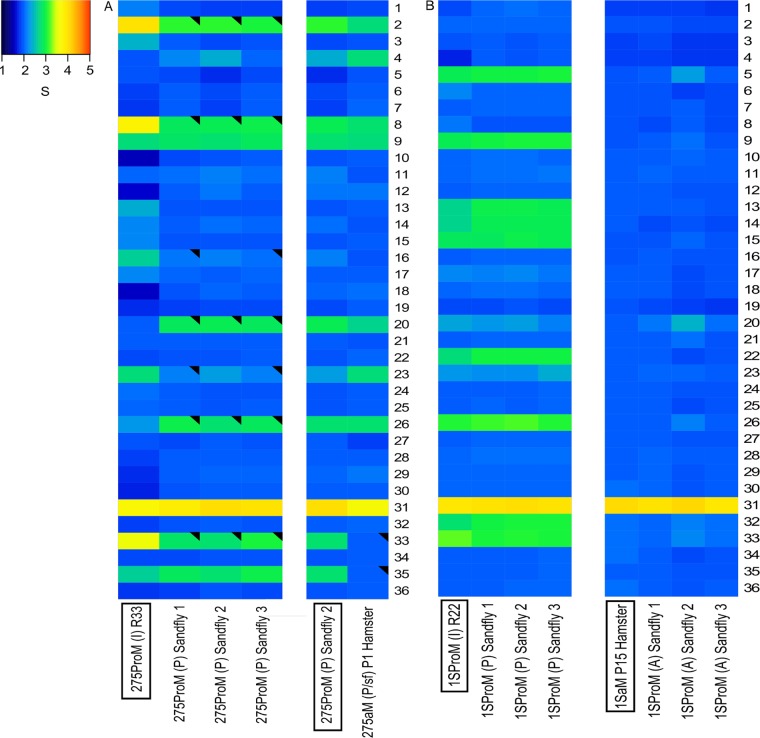
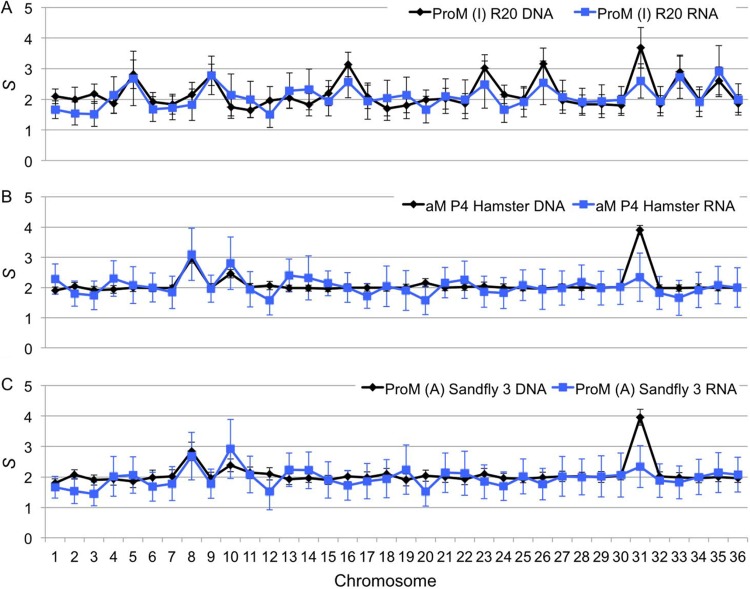
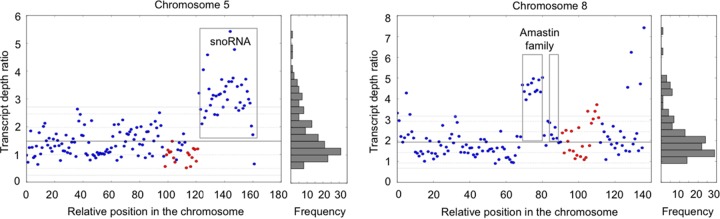
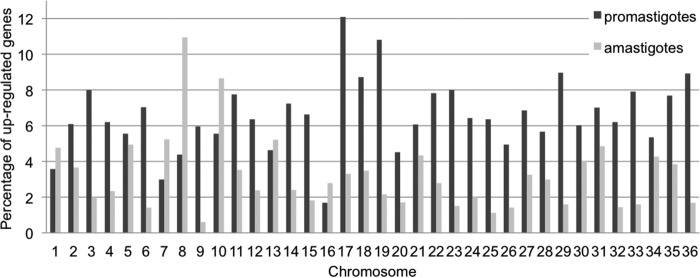
Aneuploidy is usually deleterious in multicellular organisms but appears to be tolerated and potentially beneficial in unicellular organisms, including pathogens. Leishmania, a major protozoan parasite, is emerging as a new model for aneuploidy, since in vitro-cultivated strains are highly aneuploid, with interstrain diversity and intrastrain mosaicism. The alternation of two life stages in different environments (extracellular promastigotes and intracellular amastigotes) offers a unique opportunity to study the impact of environment on aneuploidy and gene expression. We sequenced the whole genomes and transcriptomes of Leishmania donovani strains throughout their adaptation to in vivo conditions mimicking natural vertebrate and invertebrate host environments. The nucleotide sequences were almost unchanged within a strain, in contrast to highly variable aneuploidy. Although high in promastigotes in vitro, aneuploidy dropped significantly in hamster amastigotes, in a progressive and strain-specific manner, accompanied by the emergence of new polysomies. After a passage through a sand fly, smaller yet consistent karyotype changes were detected. Changes in chromosome copy numbers were correlated with the corresponding transcript levels, but additional aneuploidy-independent regulation of gene expression was observed. This affected stage-specific gene expression, downregulation of the entire chromosome 31, and upregulation of gene arrays on chromosomes 5 and 8. Aneuploidy changes in Leishmania are probably adaptive and exploited to modulate the dosage and expression of specific genes; they are well tolerated, but additional mechanisms may exist to regulate the transcript levels of other genes located on aneuploid chromosomes. Our model should allow studies of the impact of aneuploidy on molecular adaptations and cellular fitness.IMPORTANCE Aneuploidy is usually detrimental in multicellular organisms, but in several microorganisms, it can be tolerated and even beneficial. Leishmania-a protozoan parasite that kills more than 30,000 people each year-is emerging as a new model for aneuploidy studies, as unexpectedly high levels of aneuploidy are found in clinical isolates. Leishmania lacks classical regulation of transcription at initiation through promoters, so aneuploidy could represent a major adaptive strategy of this parasite to modulate gene dosage in response to stressful environments. For the first time, we document the dynamics of aneuploidy throughout the life cycle of the parasite, in vitro and in vivo We show its adaptive impact on transcription and its interaction with regulation. Besides offering a new model for aneuploidy studies, we show that further genomic studies should be done directly in clinical samples without parasite isolation and that adequate methods should be developed for this.
Keywords: Leishmania; aneuploidy; gene dosage; genomics; life cycle.
Copyright © 2017 Dumetz et al.
Figures
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
