

New Functions and Subcellular Localization Patterns of c-di-GMP Components (GGDEF Domain Proteins) in B. subtilis
- PMID: 28536559
- PMCID: PMC5422440
- DOI: 10.3389/fmicb.2017.00794
New Functions and Subcellular Localization Patterns of c-di-GMP Components (GGDEF Domain Proteins) in B. subtilis
Abstract
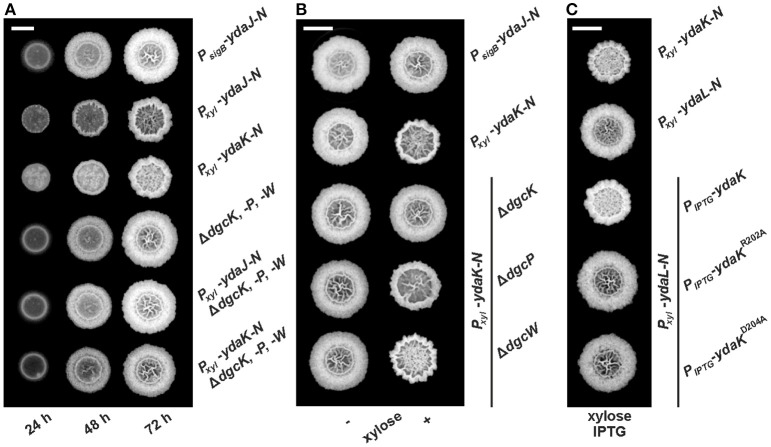
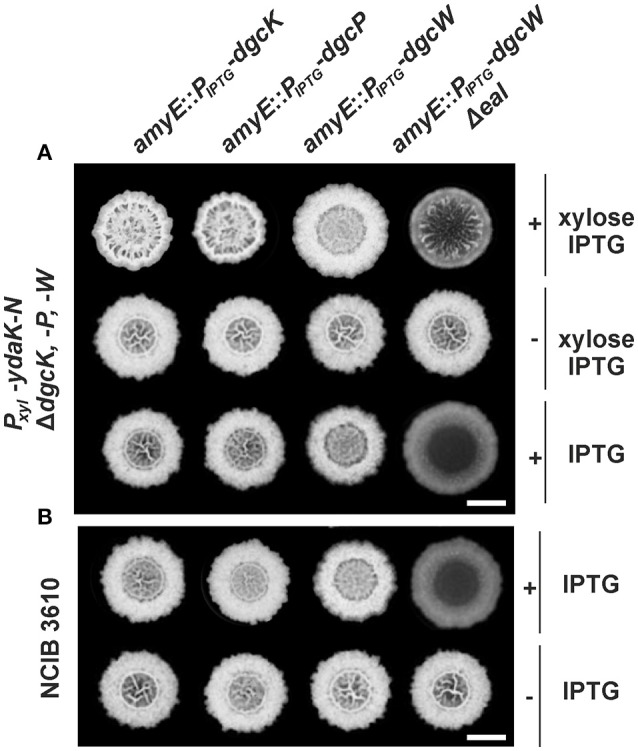
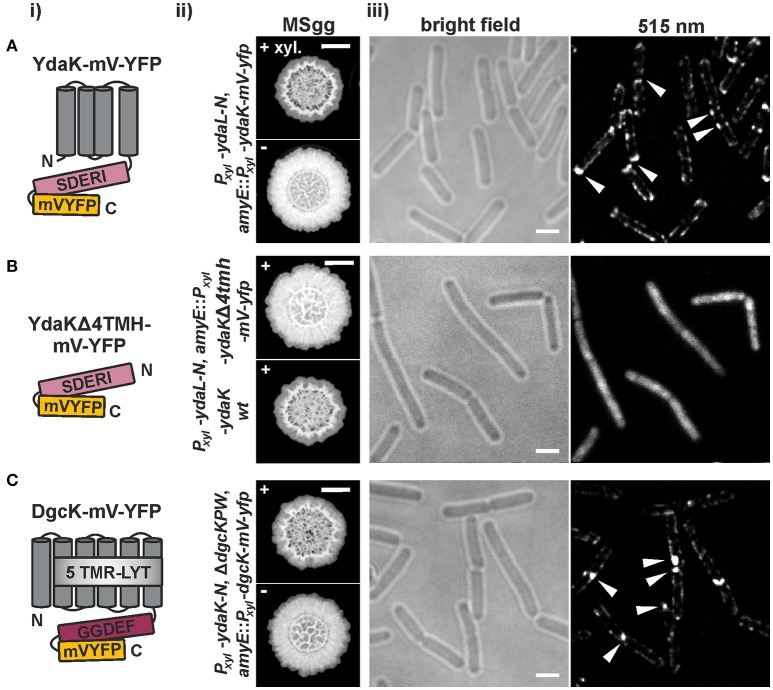
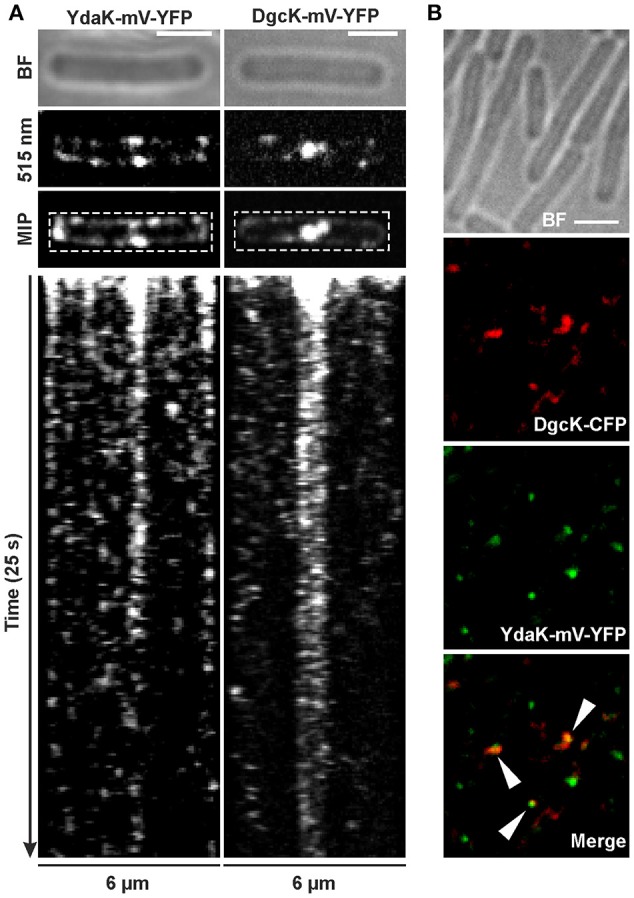
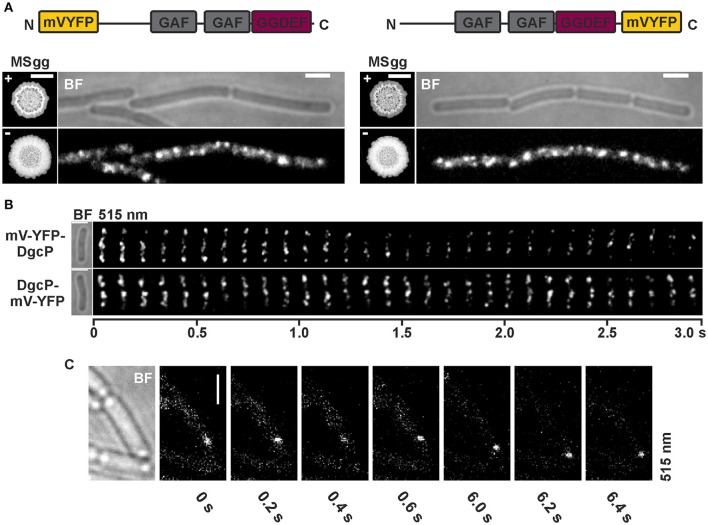
The universal and pleiotropic cyclic dinucleotide second messenger c-di-GMP is most prominently known to inversely regulate planktonic and sessile lifestyles of Gram-negative species. In the Gram-positive model organism Bacillus subtilis, intracellular c-di-GMP levels are modulated by a concise set of three diguanylate cylases (DgcK, DgcP, DgcW) and one phosphodiesterase (PdeH). Two recent studies have reported the negative influence of the c-di-GMP receptor DgrA (PilZ domain protein) on swarming motility indicating a conserved role of this second messenger across the bacterial domain. However, it has been suggested that the degenerated GGDEF protein YdaK and the inactive EAL domain protein YkuI may also function as c-di-GMP receptors regulating potentially other processes than motility. Here we describe a novel c-di-GMP dependent signaling network in B. subtilis regulating the production of an unknown exopolysaccharide (EPS) that leads to strongly altered colony morphologies upon overproduction. The network consists of the c-di-GMP receptor YdaK and the c-di-GMP synthetase DgcK. Both proteins establish a spatially close signal-effector cluster at the membrane. The cytoplasmic DgcP synthetase can complement for DgcK only upon overproduction, while the third c-di-GMP synthetase, DgcW, of B. subtilis is not part of the signaling pathway. Removal of the regulatory EAL domain from DgcW reveals a distinct function in biofilm formation. Therefore, our study is compatible with the "local pool signaling" hypothesis, but shows that in case of the yda operon, this can easily be overcome by overproduction of non-cognate DGCs, indicating that global pools can also confer signals to regulatory circuits in a Gram-positive bacterium.
Keywords: Bacillus subtilis; Biofilm formation; c-di-GMP signaling; exopolymeric substances; protein dynamics; signal transduction.
Figures
References
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases