

Sediment Resuspension and Deposition on Seagrass Leaves Impedes Internal Plant Aeration and Promotes Phytotoxic H2S Intrusion
- PMID: 28536583
- PMCID: PMC5423392
- DOI: 10.3389/fpls.2017.00657
Sediment Resuspension and Deposition on Seagrass Leaves Impedes Internal Plant Aeration and Promotes Phytotoxic H2S Intrusion
Abstract
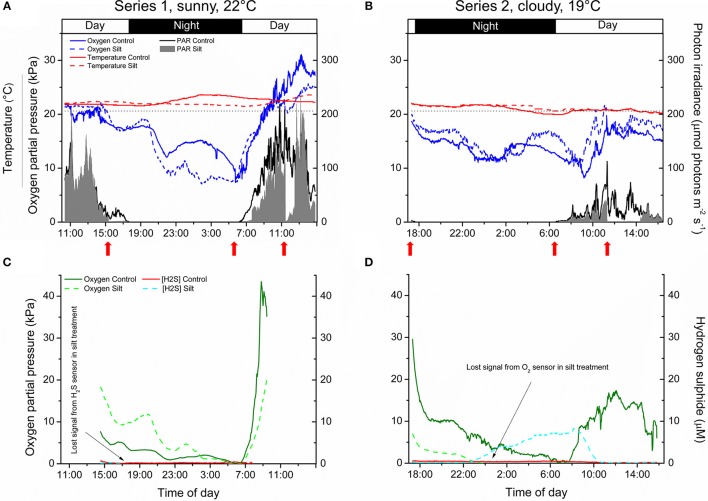
HIGHLIGHTS: Sedimentation of fine sediment particles onto seagrass leaves severely hampers the plants' performance in both light and darkness, due to inadequate internal plant aeration and intrusion of phytotoxic H2S. Anthropogenic activities leading to sediment re-suspension can have adverse effects on adjacent seagrass meadows, owing to reduced light availability and the settling of suspended particles onto seagrass leaves potentially impeding gas exchange with the surrounding water. We used microsensors to determine O2 fluxes and diffusive boundary layer (DBL) thickness on leaves of the seagrass Zostera muelleri with and without fine sediment particles, and combined these laboratory measurements with in situ microsensor measurements of tissue O2 and H2S concentrations. Net photosynthesis rates in leaves with fine sediment particles were down to ~20% of controls without particles, and the compensation photon irradiance increased from a span of 20-53 to 109-145 μmol photons m-2 s-1. An ~2.5-fold thicker DBL around leaves with fine sediment particles impeded O2 influx into the leaves during darkness. In situ leaf meristematic O2 concentrations of plants exposed to fine sediment particles were lower than in control plants and exhibited long time periods of complete meristematic anoxia during night-time. Insufficient internal aeration resulted in H2S intrusion into the leaf meristematic tissues when exposed to sediment resuspension even at relatively high night-time water-column O2 concentrations. Fine sediment particles that settle on seagrass leaves thus negatively affect internal tissue aeration and thereby the plants' resilience against H2S intrusion.
Keywords: H2S; diffusive boundary layer; dredging; in situ; microsensors; photosynthesis; seagrass; sediment.
Figures





References
-
- Barnabas A. D. (1996). Casparian band-like structures in the root hypodermis of some aquatic angiosperms. Aquat. Bot. 55, 217–225. 10.1016/S0304-3770(96)01072-8 - DOI
-
- Binzer T., Borum J., Pedersen O. (2005). Flow velocity affects internal oxygen conditions in the seagrass Cymodocea nodosa. Aquat. Bot. 83, 239–247. 10.1016/j.aquabot.2005.07.001 - DOI
-
- Borum J., Pedersen O., Greve T. M., Frankovich T. A., Zieman J. C., Fourqurean J. W., et al. (2005). The potential role of plant oxygen and sulphide dynamics in die-off events of the tropical seagrass, Thalassia testudinum. J. Ecol. 93, 148–158. 10.1111/j.1365-2745.2004.00943.x - DOI
-
- Borum J., Sand-Jensen K., Binzer T., Pedersen O., Greve T. M. (2006). Oxygen movement in seagrasses, in Seagrasses: Biology, Ecology and Conservation, eds Larkum A. W. D., Orth J. R., Duarte C. M. (Dordrecht Berlin: Springer; ), 255–270.
LinkOut - more resources
Full Text Sources
Other Literature Sources