

Understanding nucleosome dynamics and their links to gene expression and DNA replication
- PMID: 28537572
- PMCID: PMC5831138
- DOI: 10.1038/nrm.2017.47
Understanding nucleosome dynamics and their links to gene expression and DNA replication
Abstract
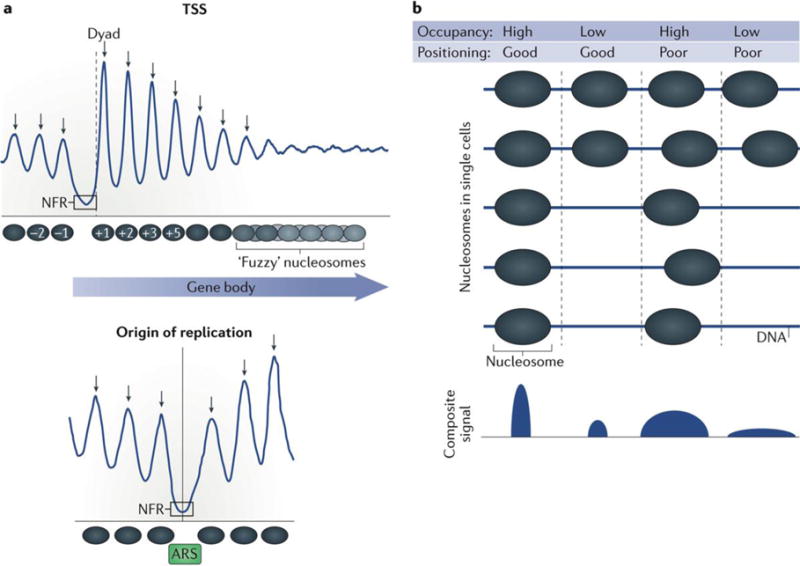
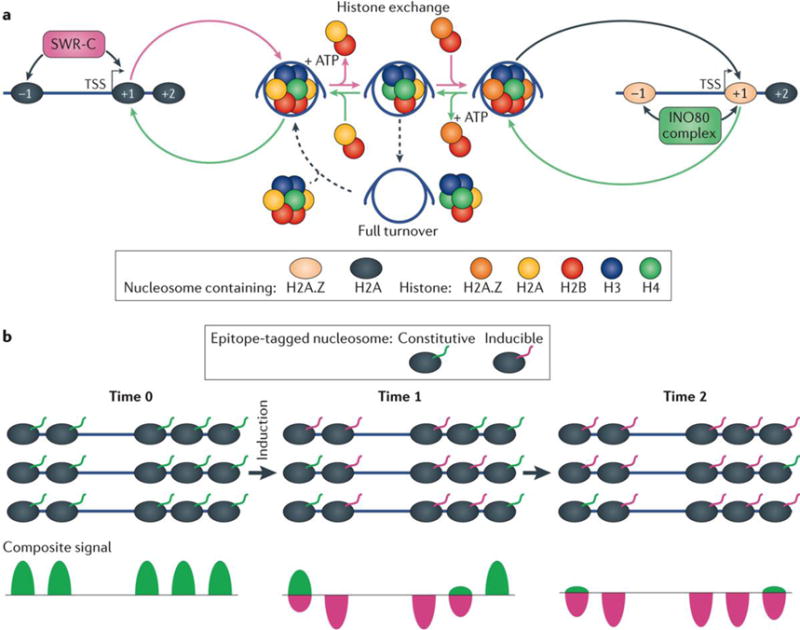
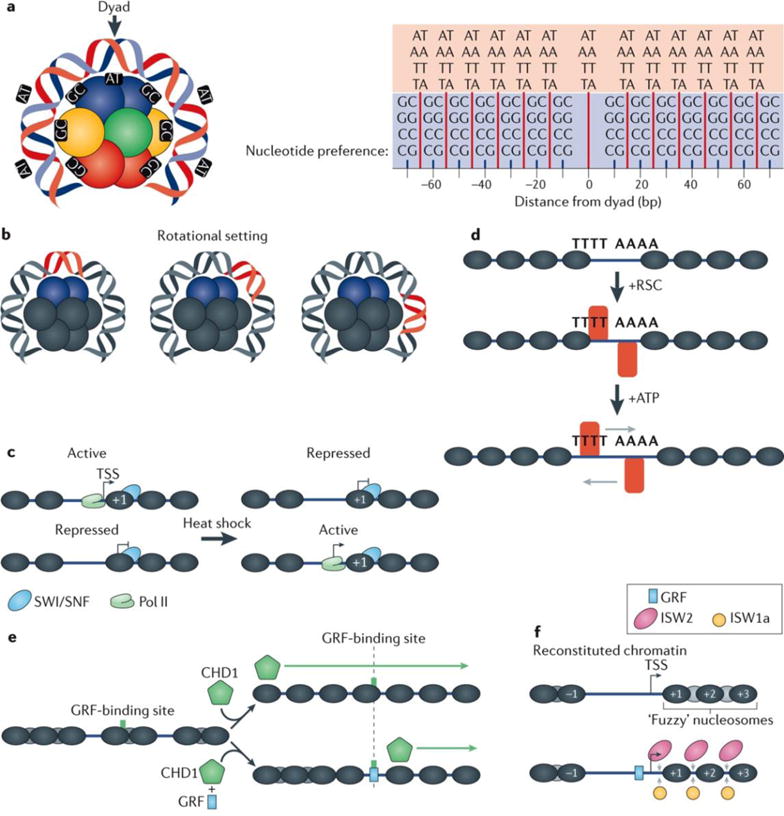
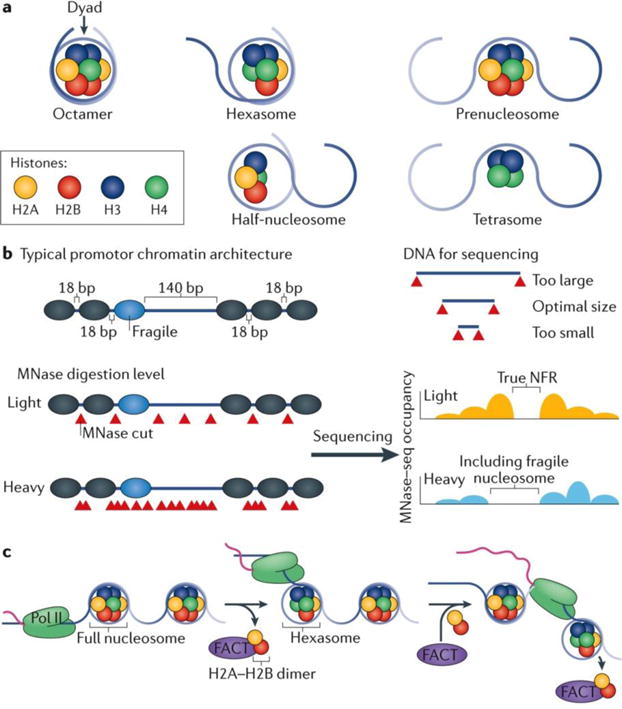
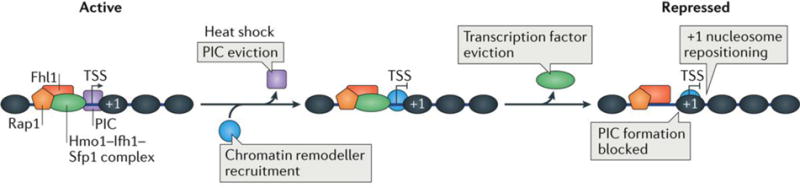
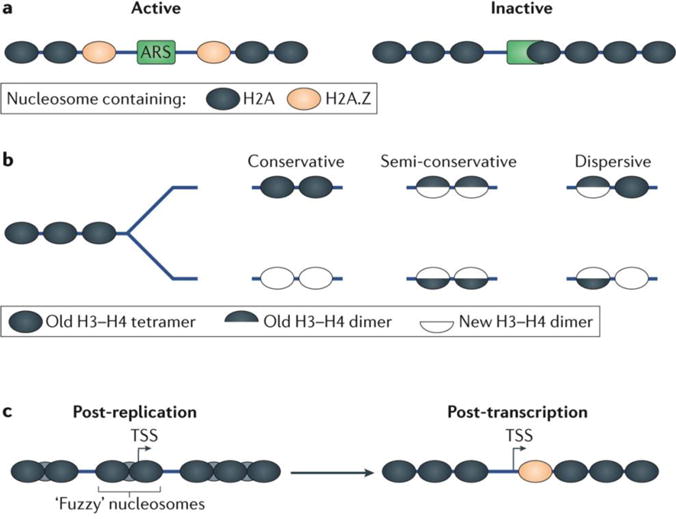
Advances in genomics technology have provided the means to probe myriad chromatin interactions at unprecedented spatial and temporal resolution. This has led to a profound understanding of nucleosome organization within the genome, revealing that nucleosomes are highly dynamic. Nucleosome dynamics are governed by a complex interplay of histone composition, histone post-translational modifications, nucleosome occupancy and positioning within chromatin, which are influenced by numerous regulatory factors, including general regulatory factors, chromatin remodellers, chaperones and polymerases. It is now known that these dynamics regulate diverse cellular processes ranging from gene transcription to DNA replication and repair.
Conflict of interest statement
The authors declare competing interests: see Web version for details.
Figures
References
-
- Luger K, Mader AW, Richmond RK, Sargent DF, Richmond TJ. Crystal structure of the nucleosome core particle at 2.8 Å resolution. Nature. 1997;389:251–260. - PubMed
-
- Kornberg RD, Lorch Y. Twenty-five years of the nucleosome, fundamental particle of the eukaryote chromosome. Cell. 1999;98:285–294. - PubMed
-
- Richmond TJ, Davey CA. The structure of DNA in the nucleosome core. Nature. 2003;423:145–150. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
