

Wolf-Hirschhorn Syndrome Candidate 1 Is Necessary for Correct Hematopoietic and B Cell Development
- PMID: 28538178
- PMCID: PMC5510986
- DOI: 10.1016/j.celrep.2017.04.069
Wolf-Hirschhorn Syndrome Candidate 1 Is Necessary for Correct Hematopoietic and B Cell Development
Abstract
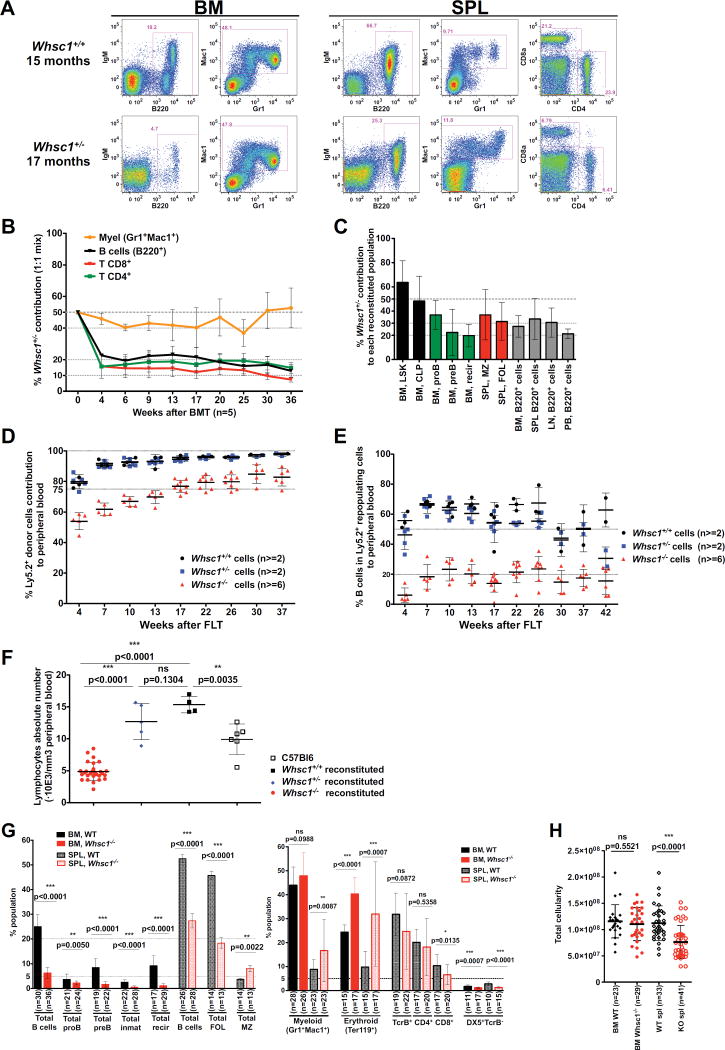
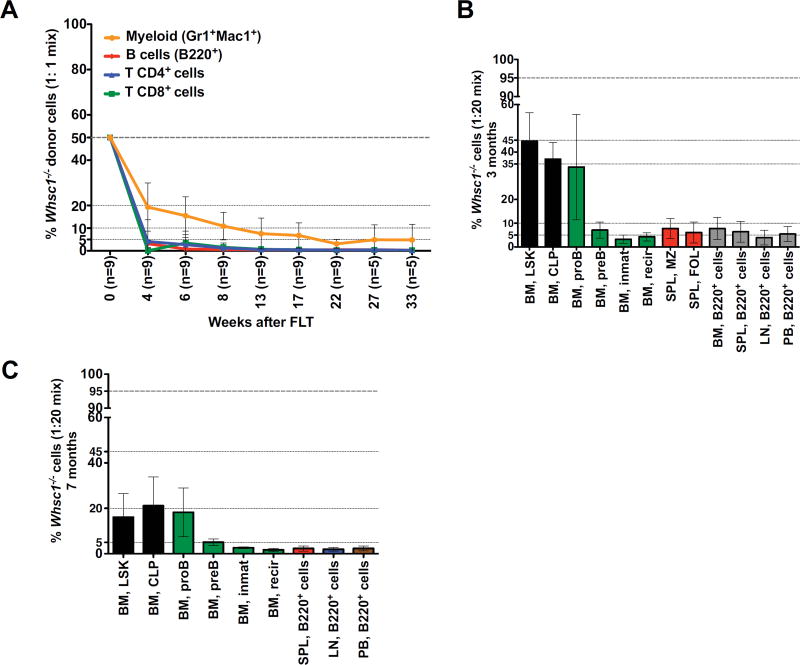
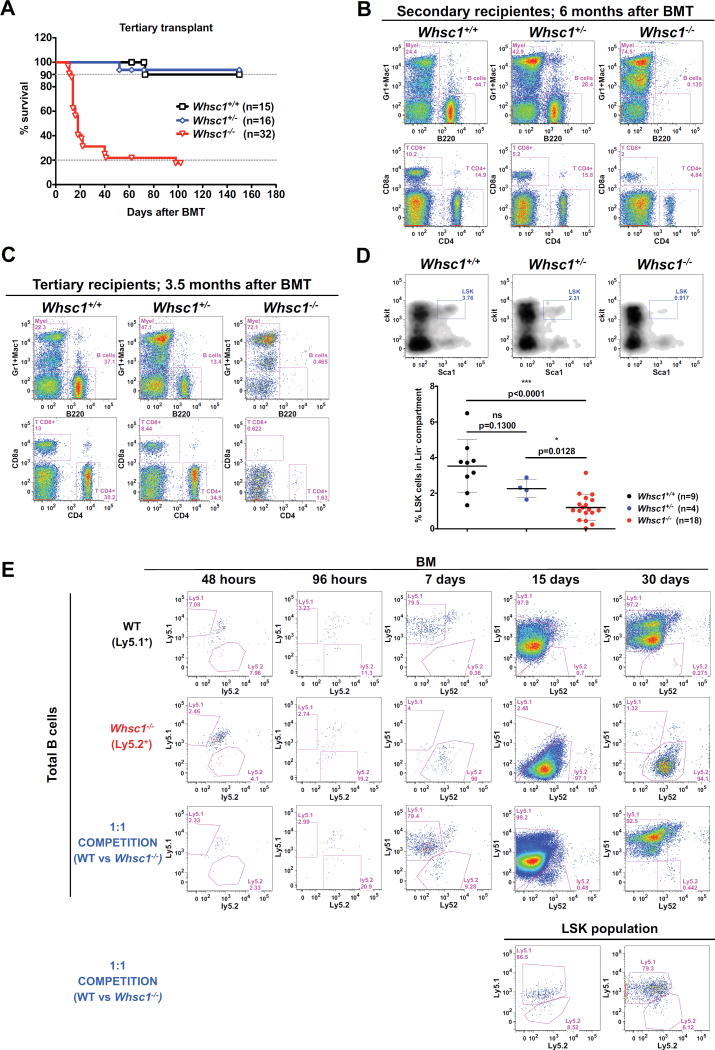
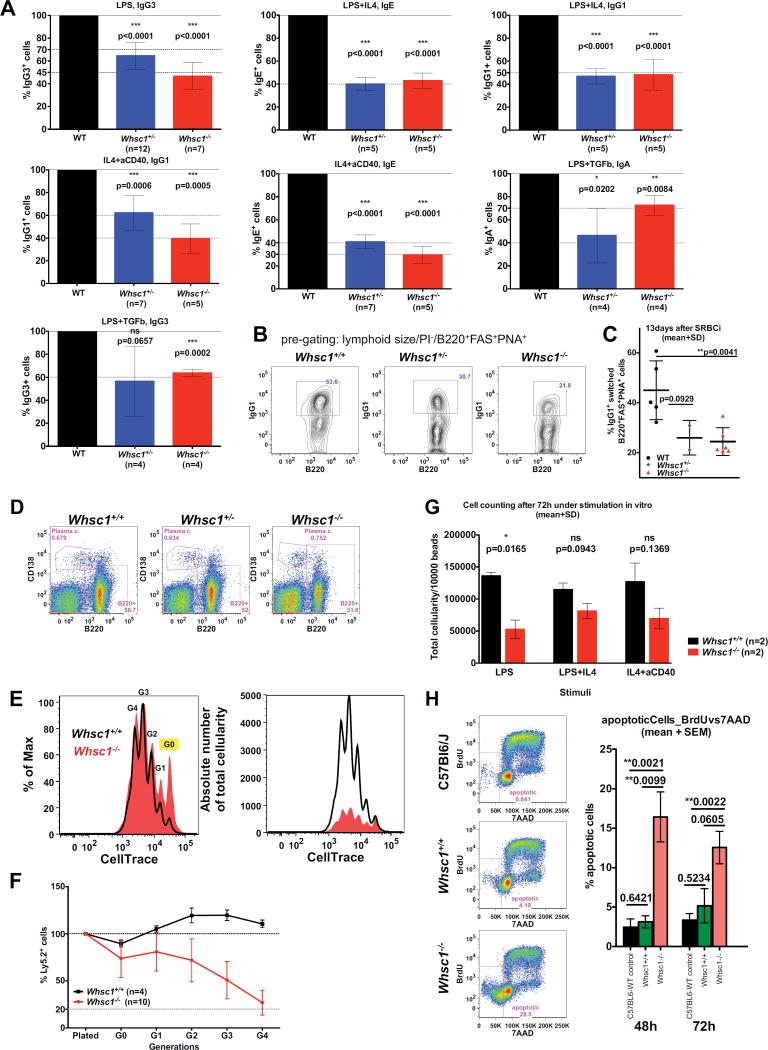
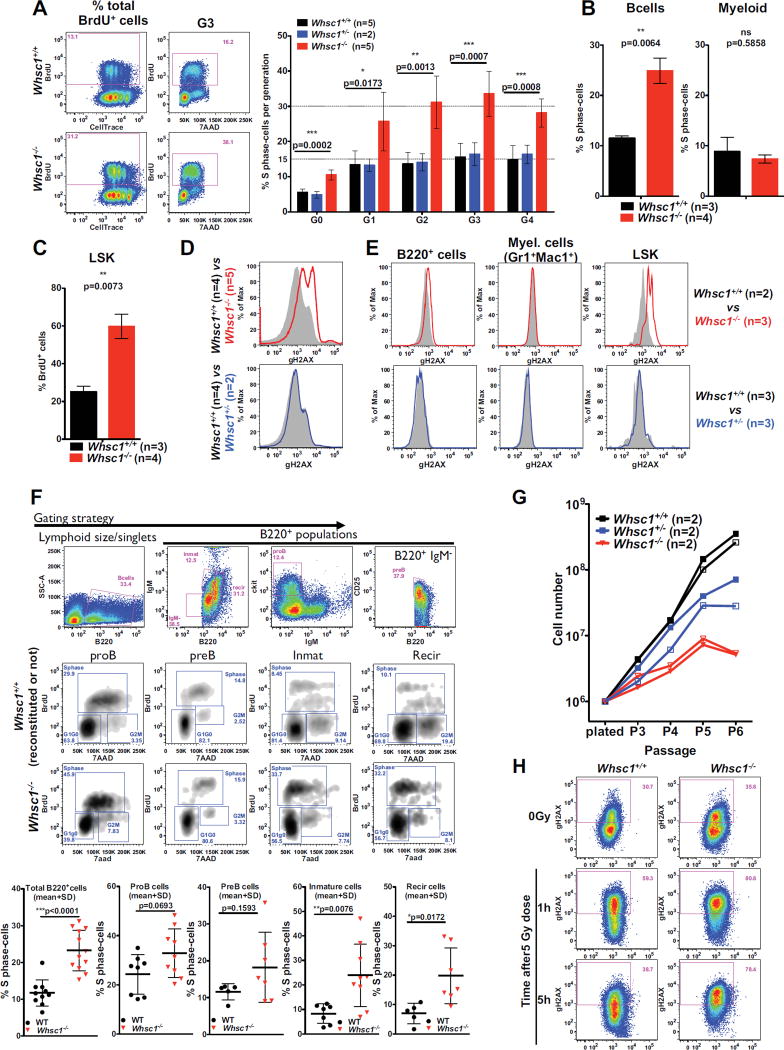
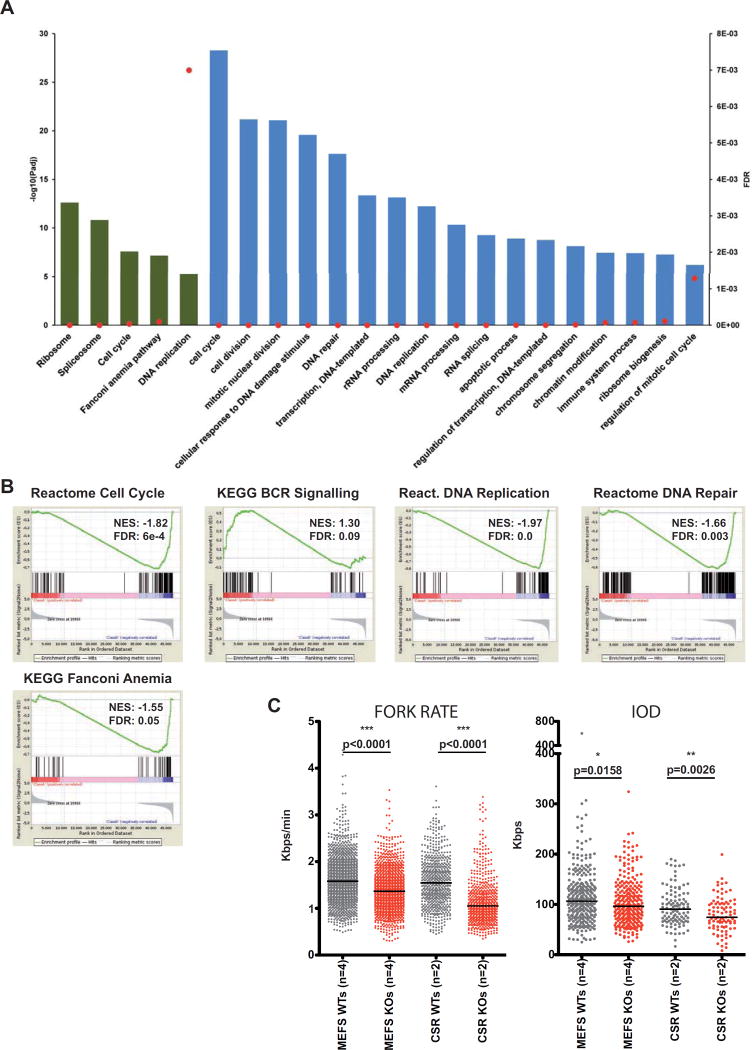
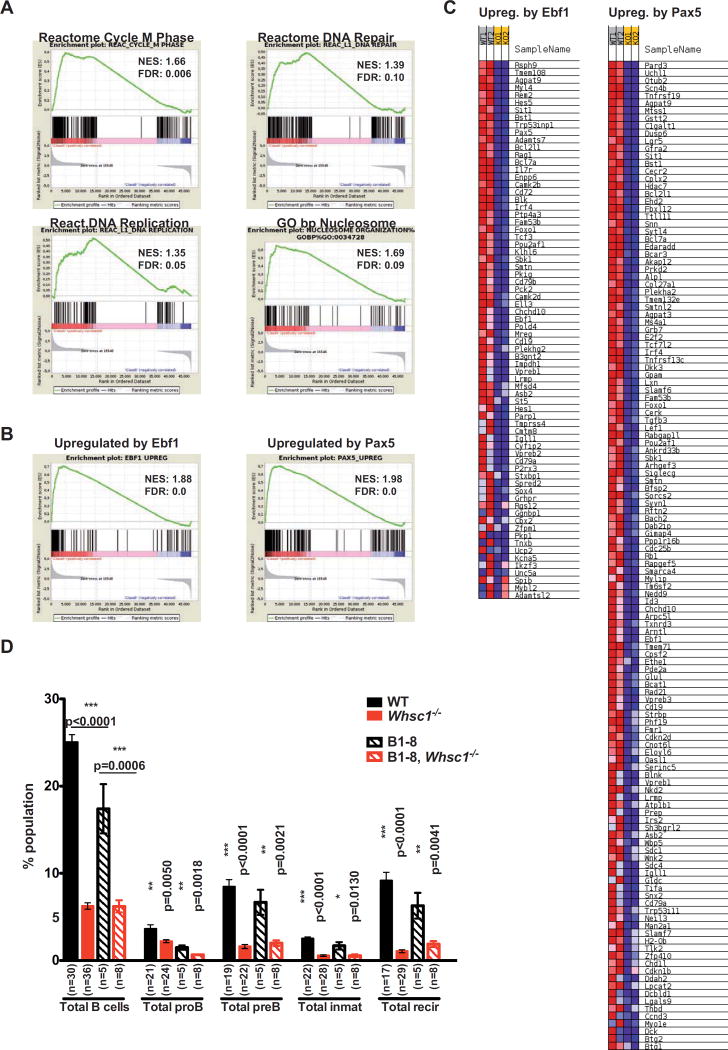
Immunodeficiency is one of the most important causes of mortality associated with Wolf-Hirschhorn syndrome (WHS), a severe rare disease originated by a deletion in chromosome 4p. The WHS candidate 1 (WHSC1) gene has been proposed as one of the main genes responsible for many of the alterations in WHS, but its mechanism of action is still unknown. Here, we present in vivo genetic evidence showing that Whsc1 plays an important role at several points of hematopoietic development. Particularly, our results demonstrate that both differentiation and function of Whsc1-deficient B cells are impaired at several key developmental stages due to profound molecular defects affecting B cell lineage specification, commitment, fitness, and proliferation, demonstrating a causal role for WHSC1 in the immunodeficiency of WHS patients.
Keywords: B cell development; H3K36 methylation; Wolf-Hirschhorn syndrome; cell cycle; class-switch recombination; hematopoiesis; hematopoietic stem cells; immunodeficiency; replicative stress.
Copyright © 2017 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
Similar articles
-
Wolf-Hirschhorn syndrome candidate 1 is involved in the cellular response to DNA damage.Proc Natl Acad Sci U S A. 2011 Aug 9;108(32):13130-4. doi: 10.1073/pnas.1110081108. Epub 2011 Jul 25. Proc Natl Acad Sci U S A. 2011. PMID: 21788515 Free PMC article.
-
Wolf-Hirschhorn Syndrome Candidate 1 (whsc1) Functions as a Tumor Suppressor by Governing Cell Differentiation.Neoplasia. 2017 Aug;19(8):606-616. doi: 10.1016/j.neo.2017.05.001. Epub 2017 Jun 24. Neoplasia. 2017. PMID: 28654864 Free PMC article.
-
A histone H3 lysine 36 trimethyltransferase links Nkx2-5 to Wolf-Hirschhorn syndrome.Nature. 2009 Jul 9;460(7252):287-91. doi: 10.1038/nature08086. Epub 2009 May 31. Nature. 2009. PMID: 19483677
-
Developmental delay and failure to thrive associated with a loss-of-function variant in WHSC1 (NSD2).Am J Med Genet A. 2018 Dec;176(12):2798-2802. doi: 10.1002/ajmg.a.40498. Epub 2018 Oct 22. Am J Med Genet A. 2018. PMID: 30345613 Review.
-
The delineation of the Wolf-Hirschhorn syndrome over six decades: Illustration of the ongoing advances in phenotype analysis and cytogenomic technology.Am J Med Genet A. 2021 Sep;185(9):2748-2755. doi: 10.1002/ajmg.a.62341. Epub 2021 May 18. Am J Med Genet A. 2021. PMID: 34002939 Review.
Cited by
-
Wolf-Hirschhorn Syndrome with Hyperparathyroidism: A Case Report and a Narrative Review of the Literature.J Pediatr Genet. 2021 Jun 26;12(4):312-317. doi: 10.1055/s-0041-1729751. eCollection 2023 Dec. J Pediatr Genet. 2021. PMID: 38162156 Free PMC article.
-
Oral Manifestations of Wolf-Hirschhorn Syndrome: Genotype-Phenotype Correlation Analysis.J Clin Med. 2020 Nov 4;9(11):3556. doi: 10.3390/jcm9113556. J Clin Med. 2020. PMID: 33158290 Free PMC article.
-
Loss of function in NSD2 causes DNA methylation signature similar to that in Wolf-Hirschhorn syndrome.Genet Med Open. 2024 Mar 14;2:101838. doi: 10.1016/j.gimo.2024.101838. eCollection 2024. Genet Med Open. 2024. PMID: 39669601 Free PMC article.
-
The role of histone methyltransferases in neurocognitive disorders associated with brain size abnormalities.Front Neurosci. 2023 Feb 10;17:989109. doi: 10.3389/fnins.2023.989109. eCollection 2023. Front Neurosci. 2023. PMID: 36845425 Free PMC article. Review.
-
The NSD2/WHSC1/MMSET methyltransferase prevents cellular senescence-associated epigenomic remodeling.Aging Cell. 2020 Jul;19(7):e13173. doi: 10.1111/acel.13173. Epub 2020 Jun 22. Aging Cell. 2020. PMID: 32573059 Free PMC article.
References
-
- Battaglia A, Carey JC, South ST. Wolf-Hirschhorn syndrome: A review and update. Am J Med Genet C Semin Med Genet. 2015;169:216–223. - PubMed
-
- Bergemann AD, Cole F, Hirschhorn K. The etiology of Wolf-Hirschhorn syndrome. Trends Genet. 2005;21:188–195. - PubMed
-
- Cobaleda C, Schebesta A, Delogu A, Busslinger M. Pax5: the guardian of B cell identity and function. Nat Immunol. 2007;8:463–470. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
