

A neural circuit architecture for angular integration in Drosophila
- PMID: 28538731
- PMCID: PMC6320684
- DOI: 10.1038/nature22343
A neural circuit architecture for angular integration in Drosophila
Abstract
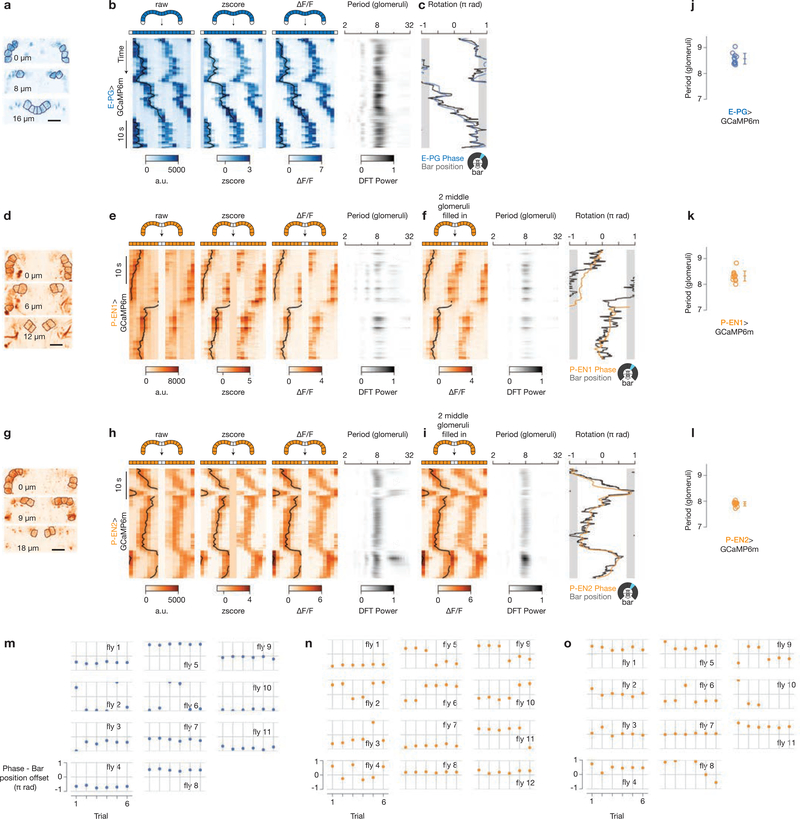
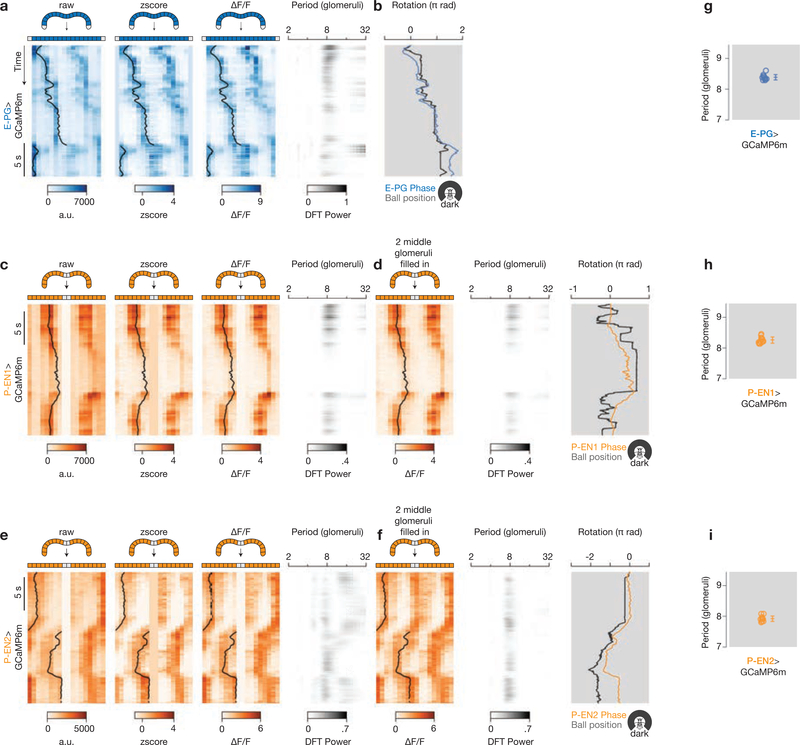
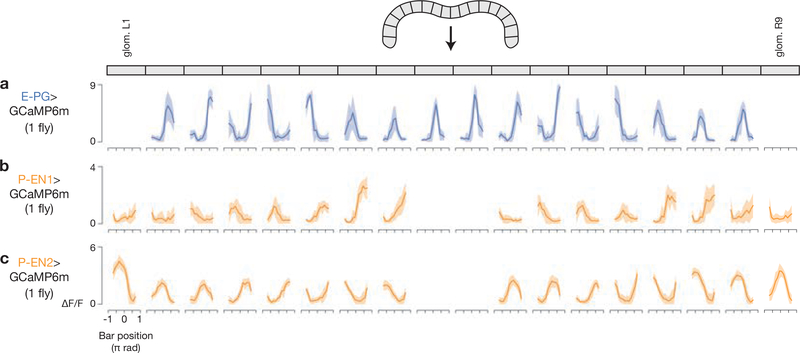
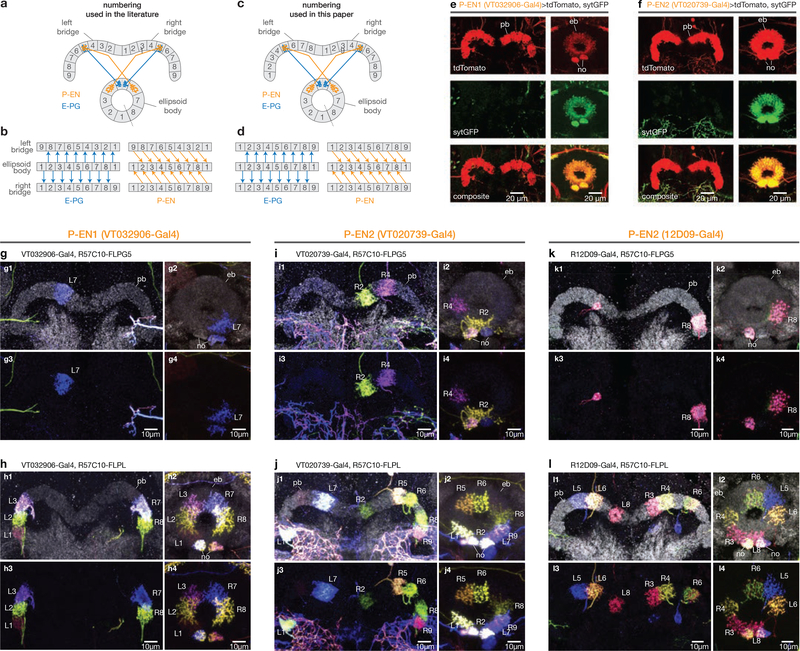
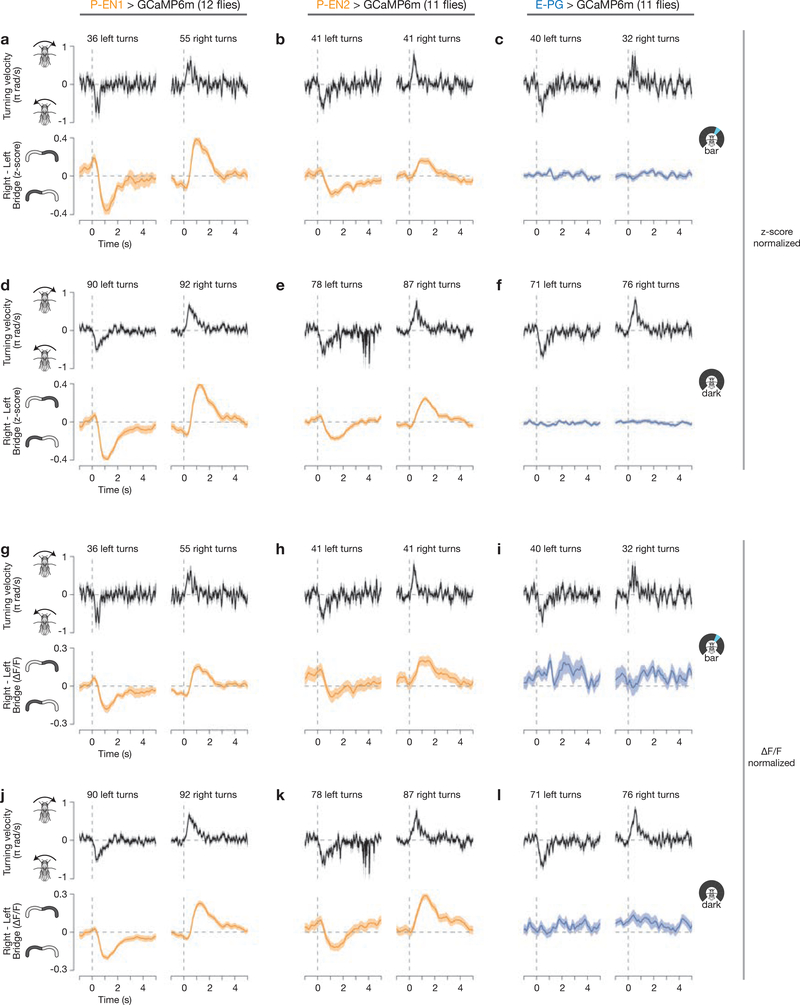
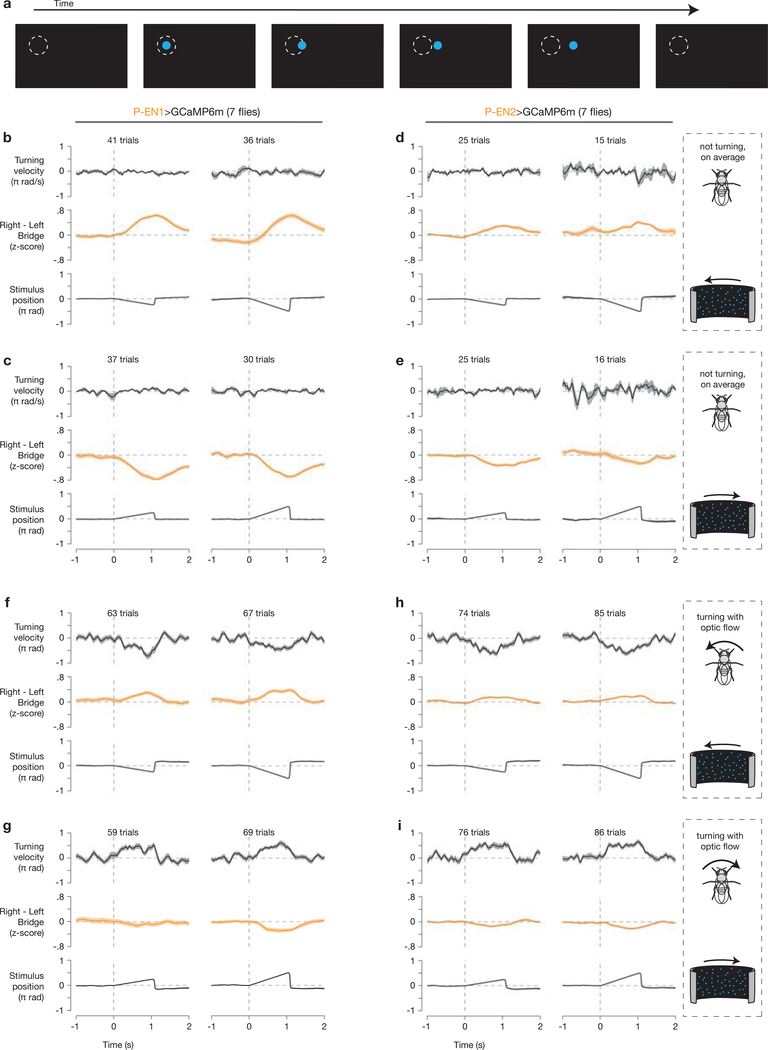
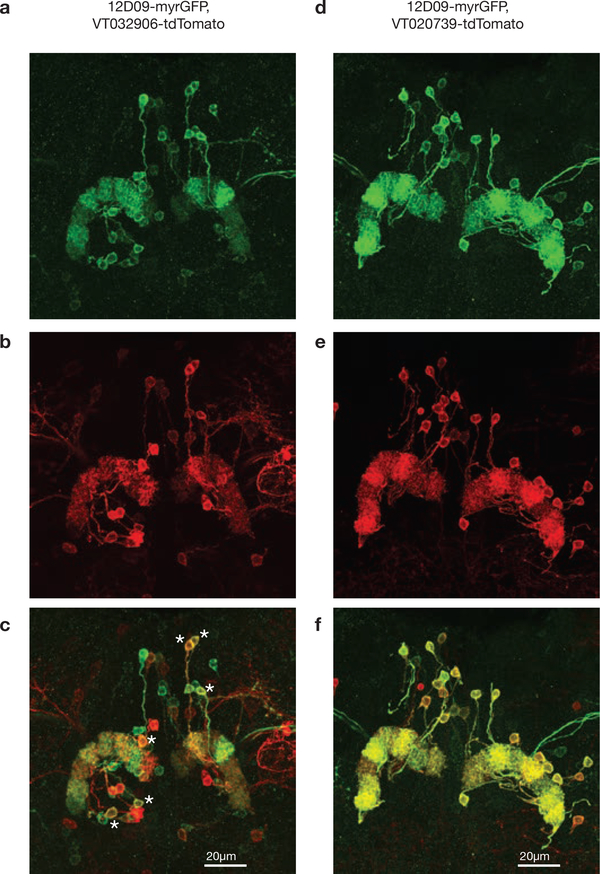
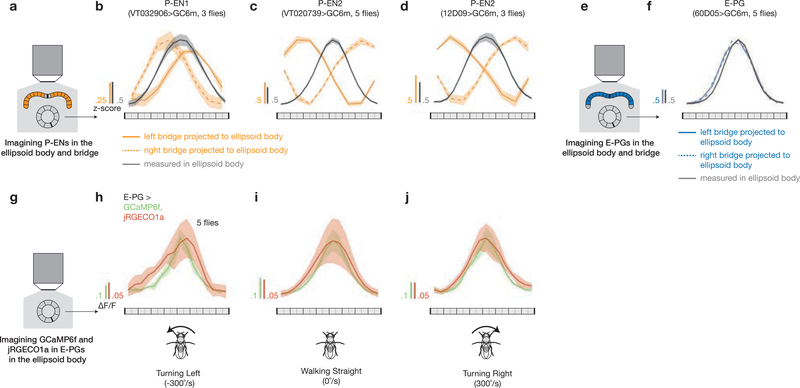
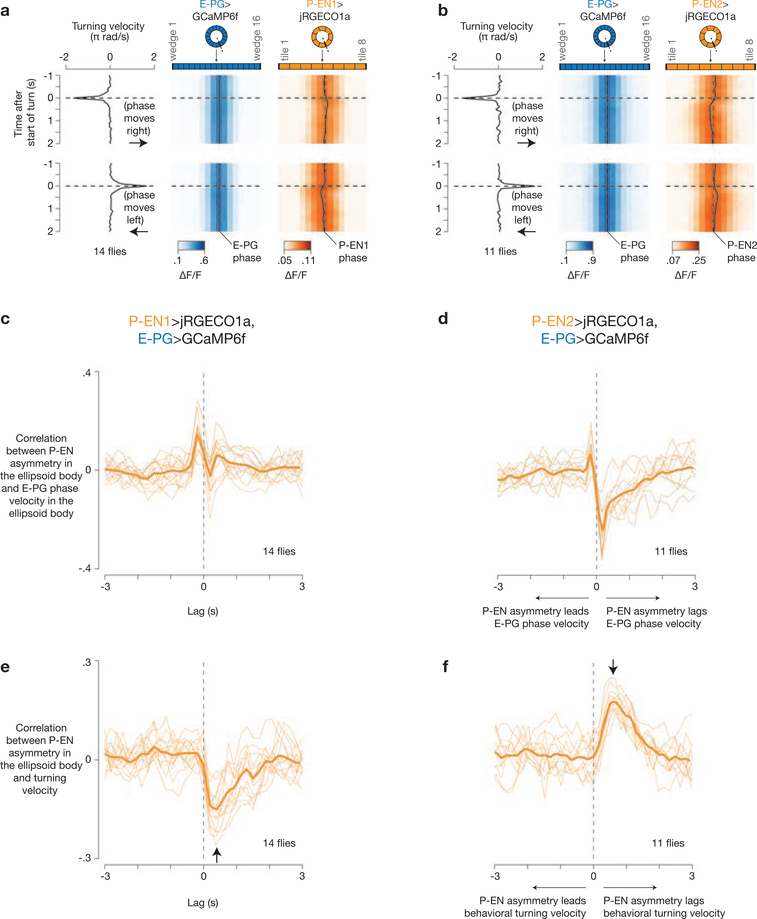
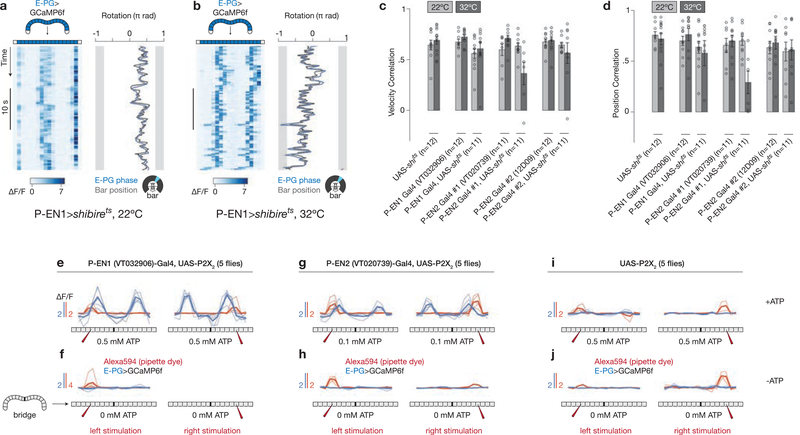
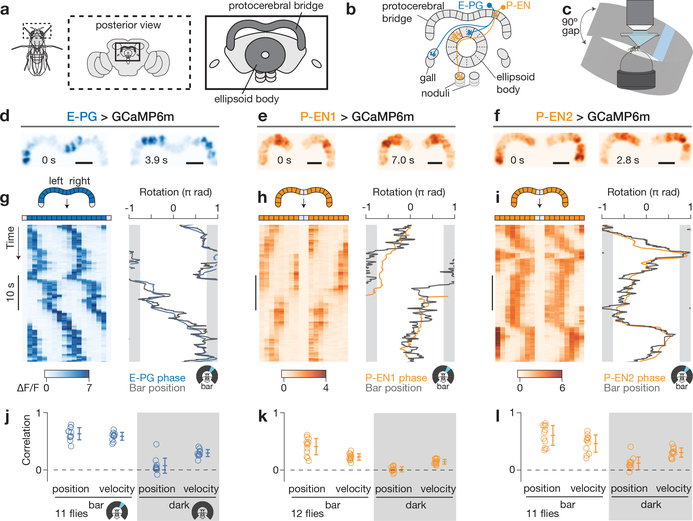
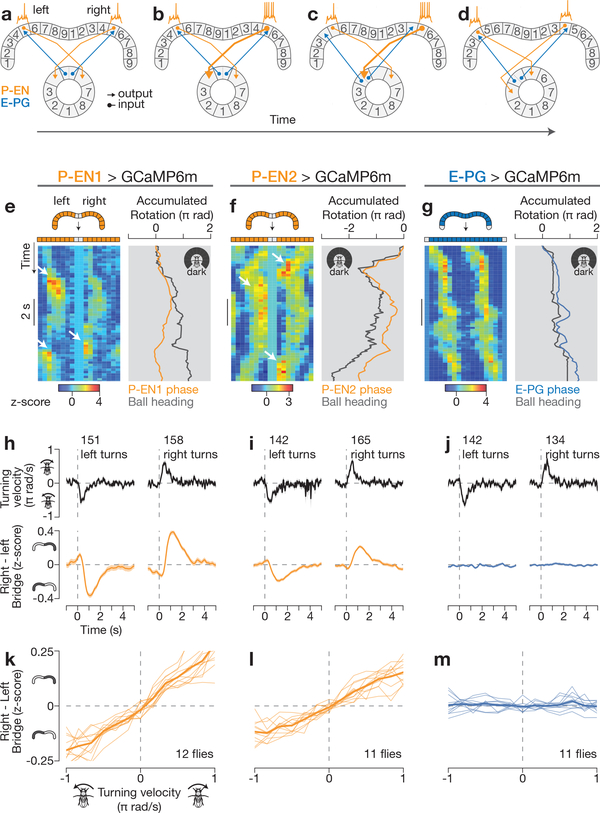
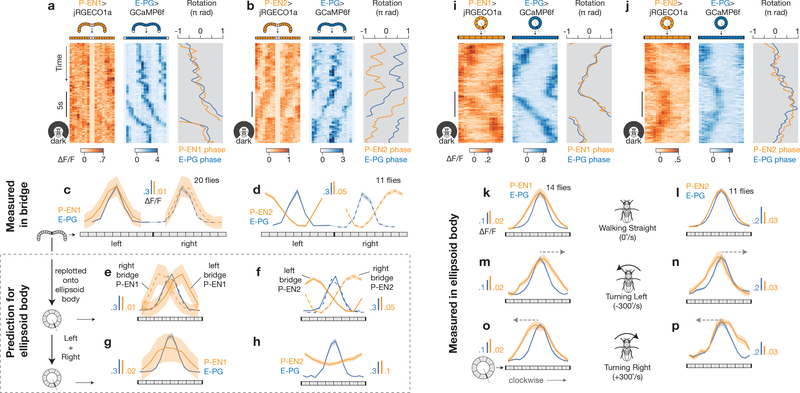
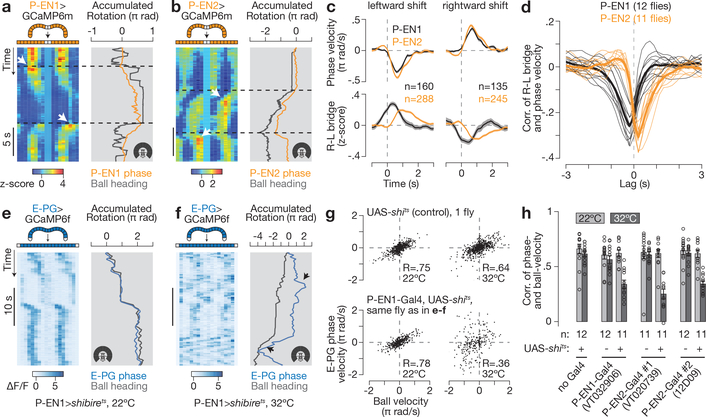
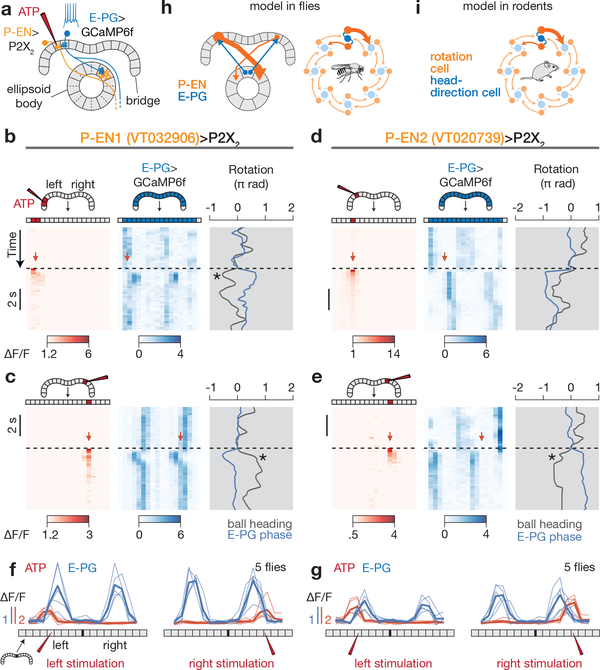
Many animals keep track of their angular heading over time while navigating through their environment. However, a neural-circuit architecture for computing heading has not been experimentally defined in any species. Here we describe a set of clockwise- and anticlockwise-shifting neurons in the Drosophila central complex whose wiring and physiology provide a means to rotate an angular heading estimate based on the fly's angular velocity. We show that each class of shifting neurons exists in two subtypes, with spatiotemporal activity profiles that suggest different roles for each subtype at the start and end of tethered-walking turns. Shifting neurons are required for the heading system to properly track the fly's heading in the dark, and stimulation of these neurons induces predictable shifts in the heading signal. The central features of this biological circuit are analogous to those of computational models proposed for head-direction cells in rodents and may shed light on how neural systems, in general, perform integration.
Figures
References
-
- O’Keefe J & Nadel L The Hippocampus as a Cognitive Map (Clarendon, 1978).
-
- Tolman EC, Ritchie BF & Kalish D Studies in spatial learning: orientation and the short-cut. J. Exp. Psychol. Gen 36, 13–24 (1946). - PubMed
-
- Etienne AS & Jeffery KJ Path integration in mammals. Hippocampus 14, 180–192 (2004). - PubMed
-
- Mittelstaedt ML & Mittelstaedt H Homing by path integration in a mammal. Naturwissenschaften 67, 566–567 (1980).
-
- Collett TS & Collett M Path integration in insects. Curr. Opin. Neurobiol 10, 757–762 (2000). - PubMed
References for Methods
-
- Fisher NI & Lee AJ A correlation coefficient for circular data. Biometrika 70, 327–332 (1983).
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
