

Microglia Are Irrelevant for Neuronal Degeneration and Axon Regeneration after Acute Injury
- PMID: 28539419
- PMCID: PMC6596505
- DOI: 10.1523/JNEUROSCI.0584-17.2017
Microglia Are Irrelevant for Neuronal Degeneration and Axon Regeneration after Acute Injury
Abstract
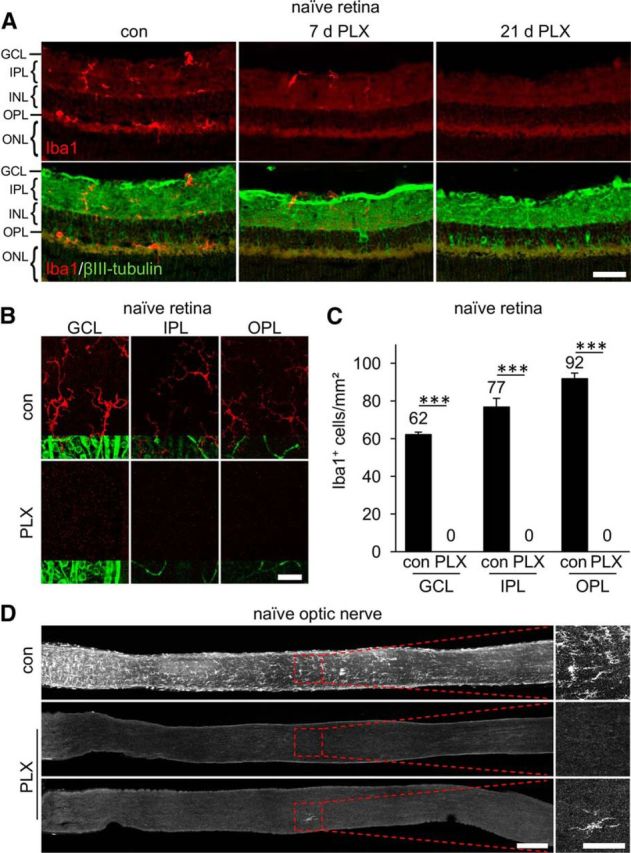
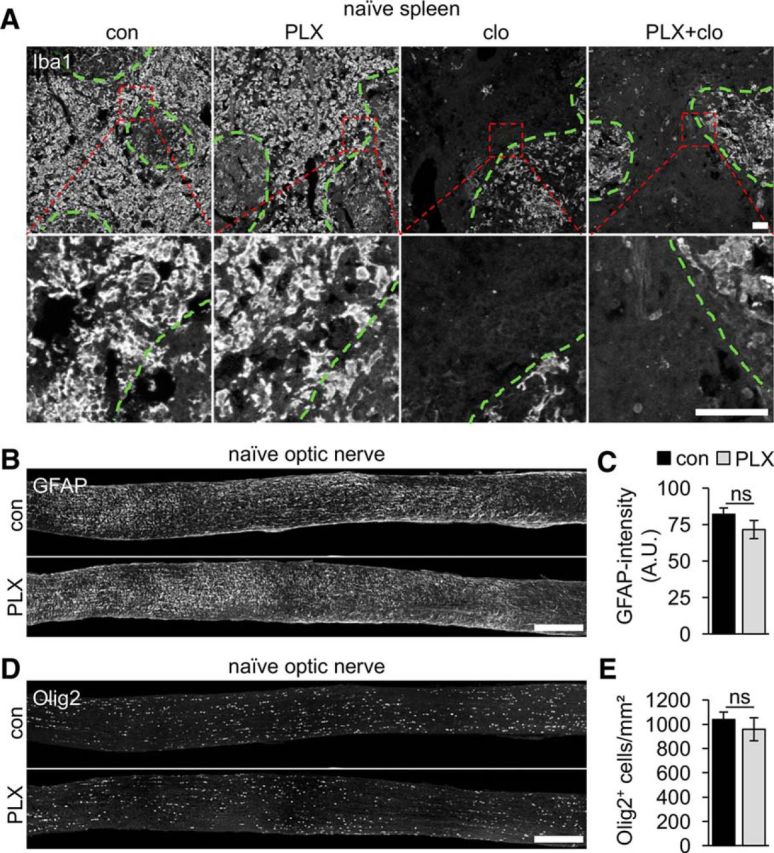
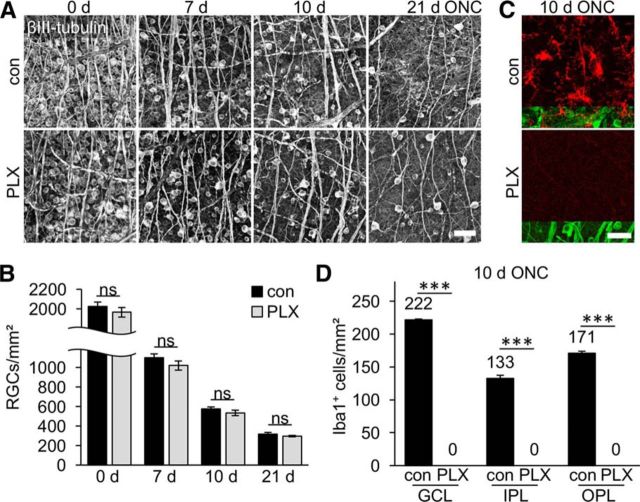
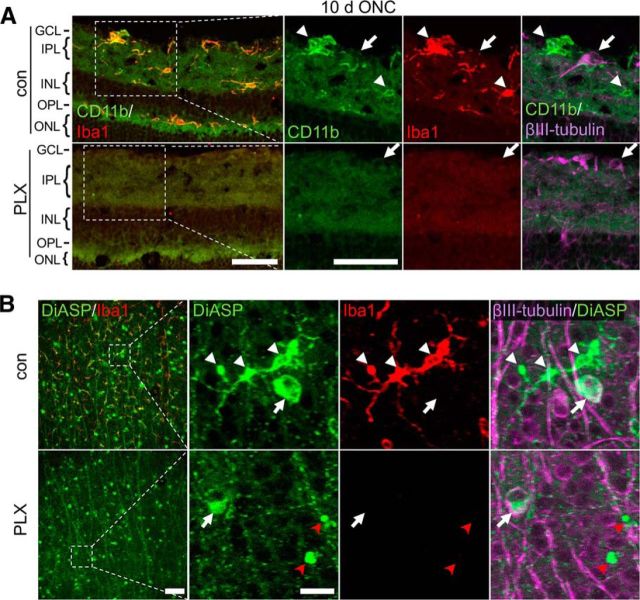
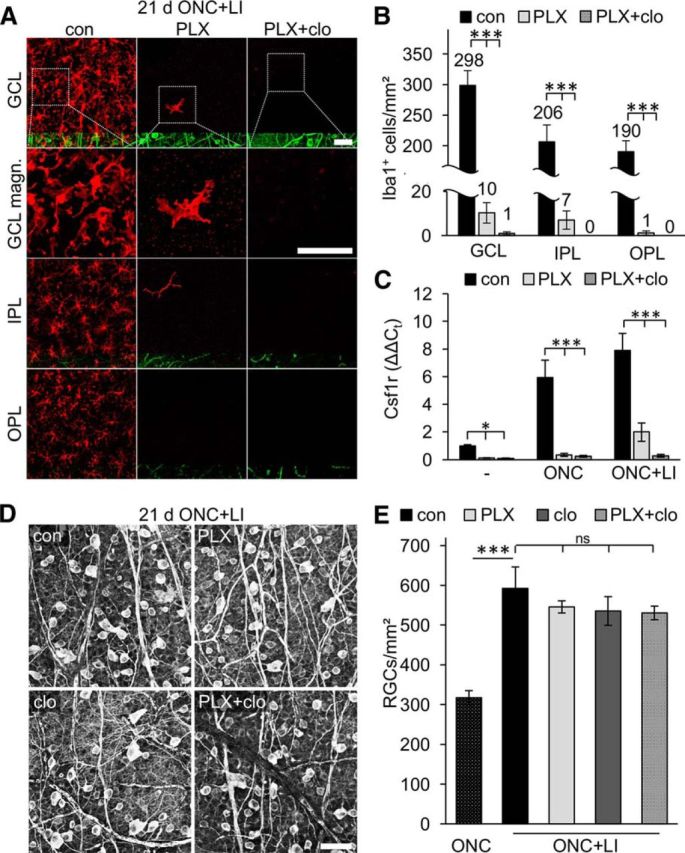
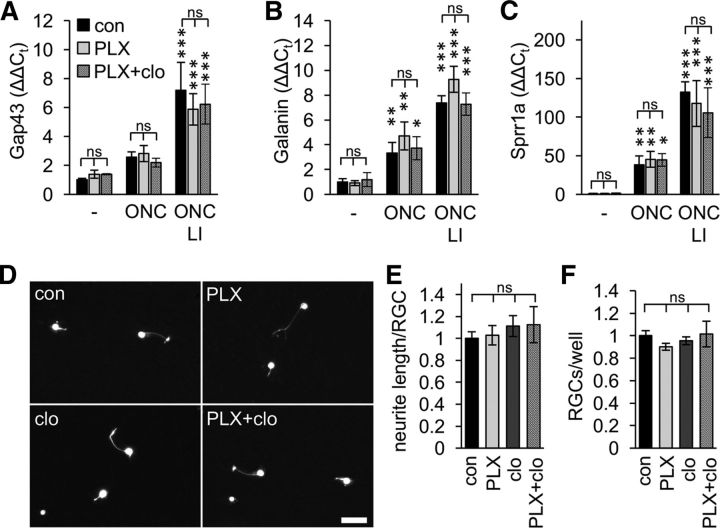
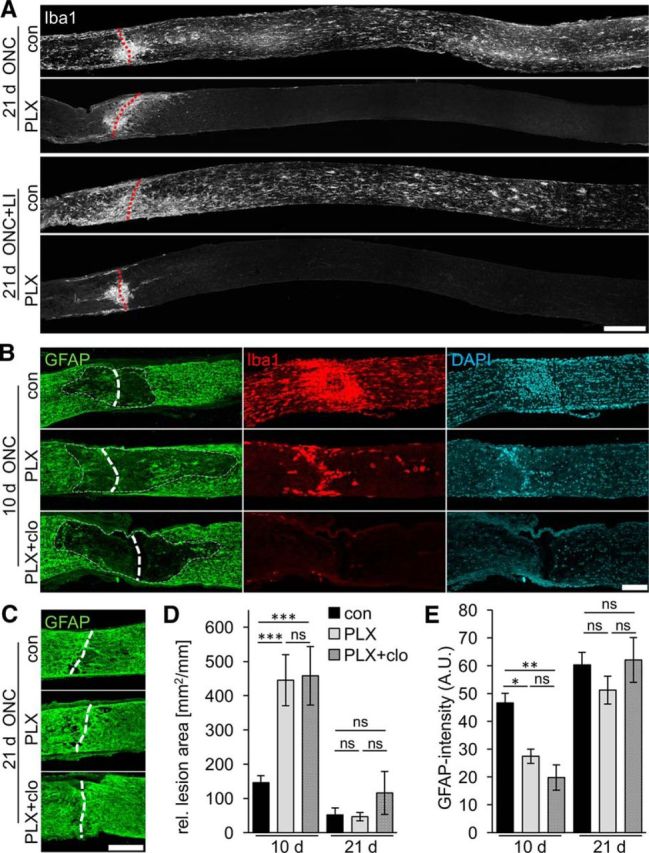
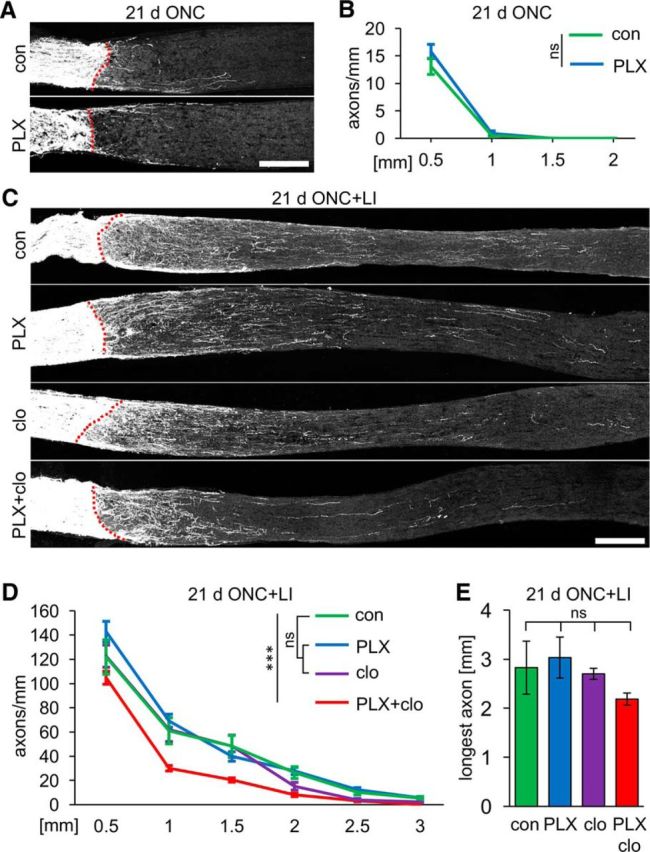
The role of microglia in degenerative and regenerative processes after damage of the nervous system remains ambiguous, partially due to the paucity of appropriate investigative methods. Here, we show that treatment with the pharmacological colony stimulating factor 1 receptor inhibitor PLX5622 specifically eliminated microglia in murine retinae and optic nerves with high efficiency. Interestingly, time course and extent of retinal ganglion cell (RGC) degeneration after optic nerve crush remained unaffected upon microglia depletion, although remnants of prelabeled apoptotic RGCs were not cleared from the retina in these animals. In addition, microglia depletion neither affected the induction of regeneration associated genes upon optic nerve injury nor the increased regenerative potential of RGCs upon lens injury (LI). However, although the repopulation of the optic nerve lesion site by astrocytes was significantly delayed upon microglia depletion, spontaneous and LI-induced axon regeneration were unaffected by PLX5622 treatment or peripheral macrophage depletion by clodronate liposome treatment. Only concurrent double depletion of microglia and infiltrated macrophages slightly, but significantly, compromised optic nerve regeneration. Therefore, microglia are not essentially involved in RGC degeneration or axonal regeneration after acute CNS injury.SIGNIFICANCE STATEMENT The roles of microglia, the phagocytosing cells of the CNS, and invading macrophages in degenerative and regenerative processes after injury are still controversial and insufficiently characterized. Here, we show that application of a CSF1R inhibitor eliminated virtually all microglia from the visual system, whereas macrophages were spared. Specific microglia depletion impaired the removal of dead labeled retinal ganglion cells after optic nerve crush, but remarkable had no influence on their degeneration. Similarly, optic nerve regeneration was completely unaffected, although repopulation of the lesion site by astrocytes was delayed significantly. Therefore, contrary to previous reports, this experimental approach revealed that microglia seemingly neither promote nor inhibit neuronal degeneration or axonal regrowth within the injured visual system.
Keywords: apoptosis; axon regeneration; lens injury; microglia; neuroprotection; optic nerve; retina.
Copyright © 2017 the authors 0270-6474/17/376113-12$15.00/0.
Figures
References
-
- Bennett ML, Bennett FC, Liddelow SA, Ajami B, Zamanian JL, Fernhoff NB, Mulinyawe SB, Bohlen CJ, Adil A, Tucker A, Weissman IL, Chang EF, Li G, Grant GA, Hayden Gephart MG, Barres BA (2016) New tools for studying microglia in the mouse and human CNS. Proc Natl Acad Sci U S A 113:E1738–E1746. 10.1073/pnas.1525528113 - DOI - PMC - PubMed
-
- Bosco A, Inman DM, Steele MR, Wu G, Soto I, Marsh-Armstrong N, Hubbard WC, Calkins DJ, Horner PJ, Vetter ML (2008) Reduced retina microglial activation and improved optic nerve integrity with minocycline treatment in the DBA/2J mouse model of glaucoma. Invest Ophthalmol Vis Sci 49:1437–1446. 10.1167/iovs.07-1337 - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous