

Bolstering the Number and Function of HSV-1-Specific CD8+ Effector Memory T Cells and Tissue-Resident Memory T Cells in Latently Infected Trigeminal Ganglia Reduces Recurrent Ocular Herpes Infection and Disease
- PMID: 28539429
- PMCID: PMC5515716
- DOI: 10.4049/jimmunol.1700145
Bolstering the Number and Function of HSV-1-Specific CD8+ Effector Memory T Cells and Tissue-Resident Memory T Cells in Latently Infected Trigeminal Ganglia Reduces Recurrent Ocular Herpes Infection and Disease
Abstract
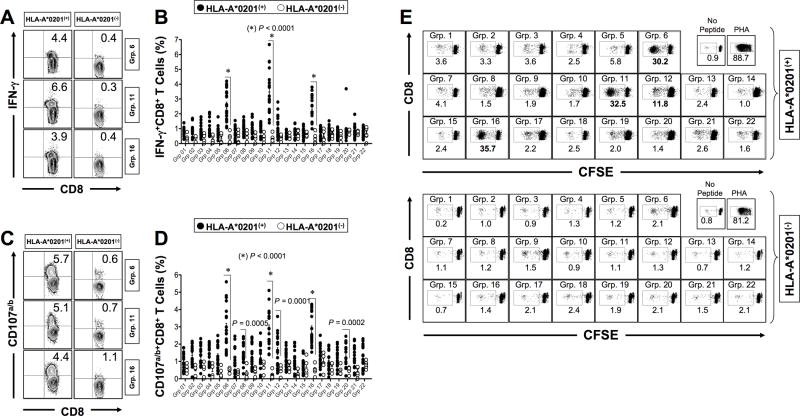
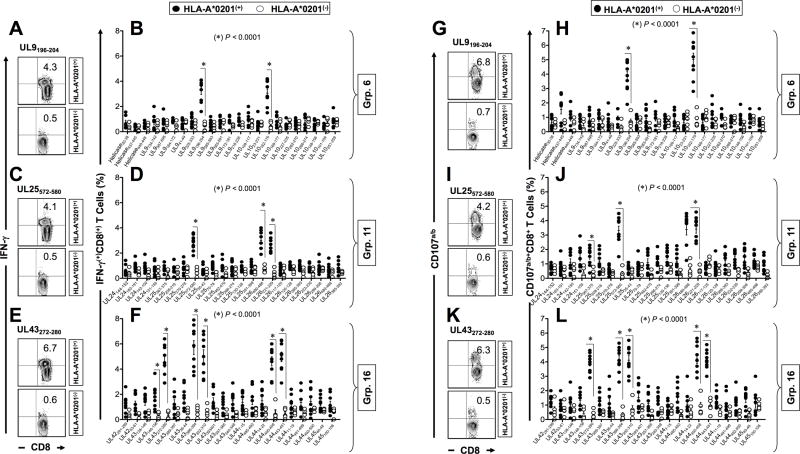
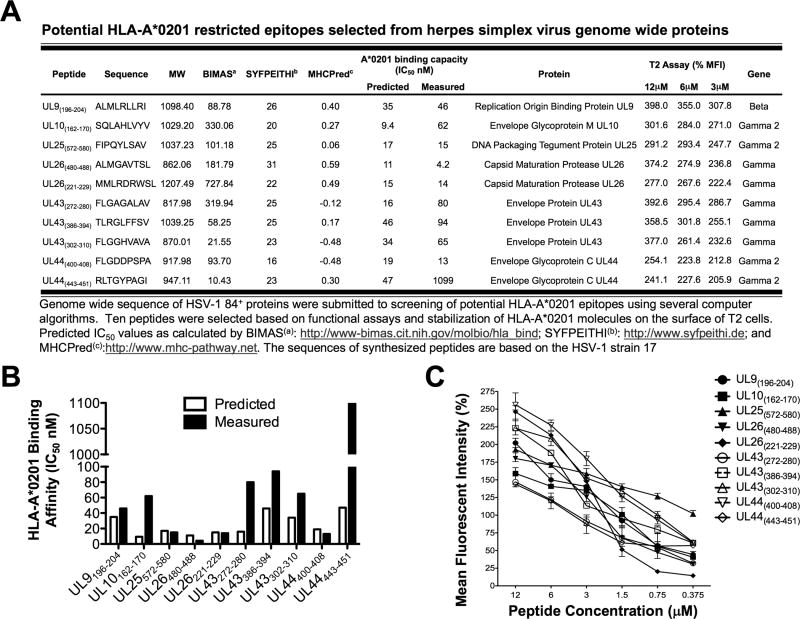
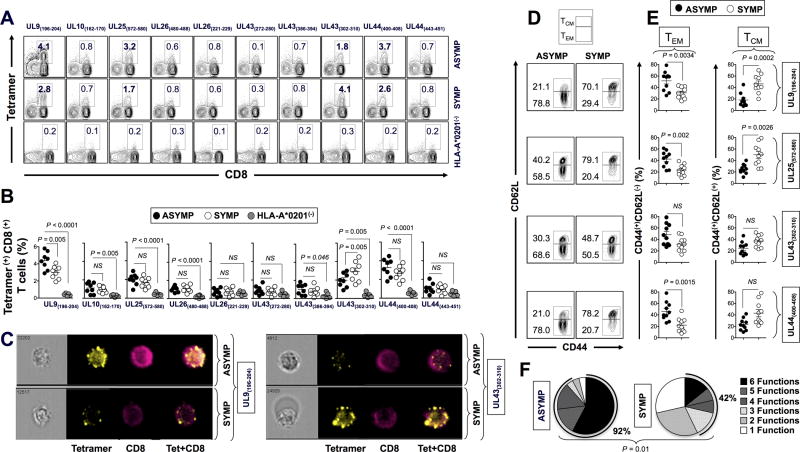
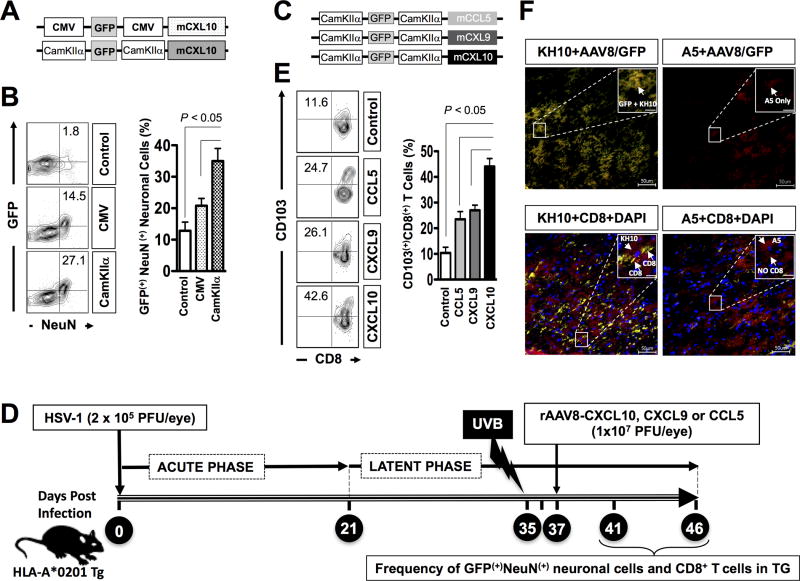
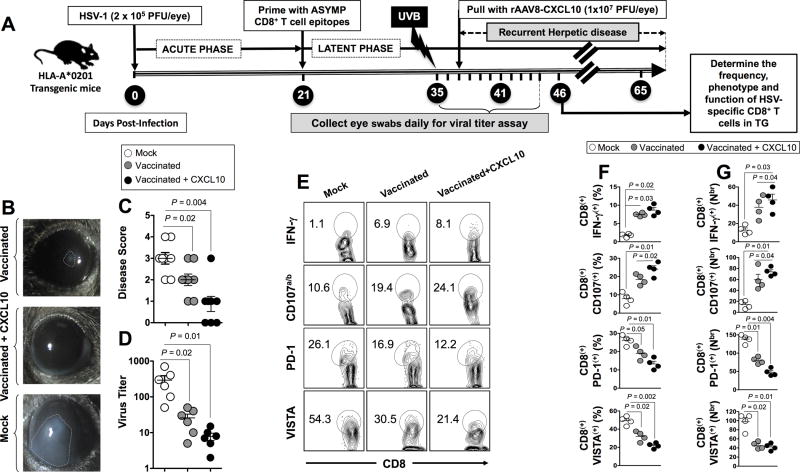
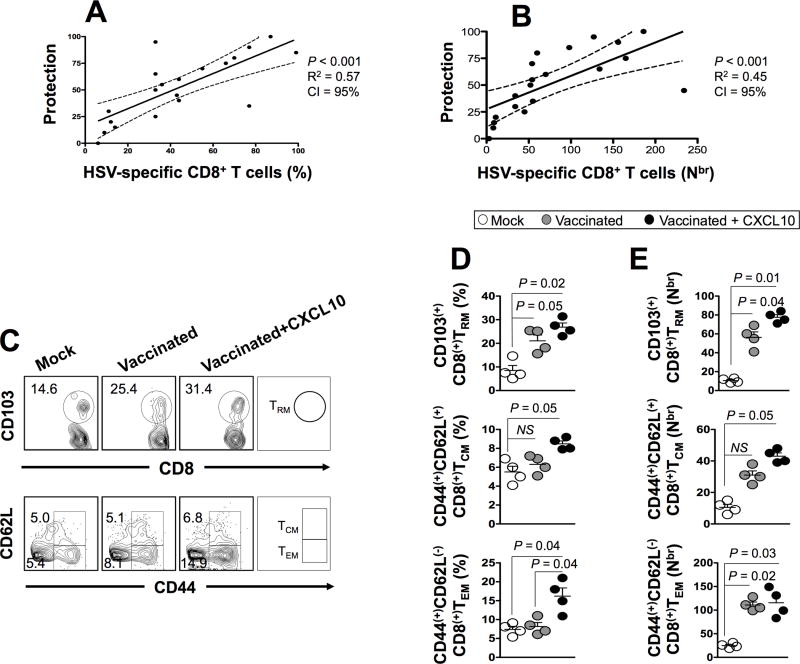
HSV type 1 (HSV-1) is a prevalent human pathogen that infects >3.72 billion individuals worldwide and can cause potentially blinding recurrent corneal herpetic disease. HSV-1 establishes latency within sensory neurons of trigeminal ganglia (TG), and TG-resident CD8+ T cells play a critical role in preventing its reactivation. The repertoire, phenotype, and function of protective CD8+ T cells are unknown. Bolstering the apparent feeble numbers of CD8+ T cells in TG remains a challenge for immunotherapeutic strategies. In this study, a comprehensive panel of 467 HLA-A*0201-restricted CD8+ T cell epitopes was predicted from the entire HSV-1 genome. CD8+ T cell responses to these genome-wide epitopes were compared in HSV-1-seropositive symptomatic individuals (with a history of numerous episodes of recurrent herpetic disease) and asymptomatic (ASYMP) individuals (who are infected but never experienced any recurrent herpetic disease). Frequent polyfunctional HSV-specific IFN-γ+CD107a/b+CD44highCD62LlowCD8+ effector memory T cells were detected in ASYMP individuals and were primarily directed against three "ASYMP" epitopes. In contrast, symptomatic individuals have more monofunctional CD44highCD62LhighCD8+ central memory T cells. Furthermore, therapeutic immunization with an innovative prime/pull vaccine, based on priming with multiple ASYMP epitopes (prime) and neurotropic TG delivery of the T cell-attracting chemokine CXCL10 (pull), boosted the number and function of CD44highCD62LlowCD8+ effector memory T cells and CD103highCD8+ tissue-resident T cells in TG of latently infected HLA-A*0201-transgenic mice and reduced recurrent ocular herpes following UV-B-induced reactivation. These findings have profound implications in the development of T cell-based immunotherapeutic strategies to treat blinding recurrent herpes infection and disease.
Copyright © 2017 by The American Association of Immunologists, Inc.
Conflict of interest statement
Conflict of interest: The authors have declared that no conflict of interest exists
Figures
References
-
- Srivastava R, Khan AA, Garg S, Syed SA, Furness JN, Vahed H, Pham T, Yu HT, Nesburn AB, BenMohamed L. Human Asymptomatic Epitopes Identified from the Herpes Simplex Virus Tegument Protein VP13/14 (UL47) Preferentially Recall Polyfunctional Effector Memory CD44highCD62LlowCD8+ TEM Cells and Protect "Humanized" HLA-A*02:01 Transgenic Mice Against Ocular Herpes. J Virol 2016 - PMC - PubMed
-
- Samandary S, Kridane-Miledi H, Sandoval JS, Choudhury Z, Langa-Vives F, Spencer D, Chentoufi AA, Lemonnier FA, BenMohamed L. Associations of HLA-A, HLA-B and HLA-C alleles frequency with prevalence of herpes simplex virus infections and diseases across global populations: implication for the development of an universal CD8+ T-cell epitope-based vaccine. Hum Immunol. 2014;75:715–729. - PMC - PubMed
-
- Zhang X, Dervillez X, Chentoufi AA, Badakhshan T, Bettahi I, Benmohamed L. Targeting the genital tract mucosa with a lipopeptide/recombinant adenovirus prime/boost vaccine induces potent and long-lasting CD8+ T cell immunity against herpes: importance of MyD88. J Immunol. 2012;189:4496–4509. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
