

Estrogens in Male Physiology
- PMID: 28539434
- PMCID: PMC6151497
- DOI: 10.1152/physrev.00018.2016
Estrogens in Male Physiology
Abstract
Estrogens have historically been associated with female reproduction, but work over the last two decades established that estrogens and their main nuclear receptors (ESR1 and ESR2) and G protein-coupled estrogen receptor (GPER) also regulate male reproductive and nonreproductive organs. 17β-Estradiol (E2) is measureable in blood of men and males of other species, but in rete testis fluids, E2 reaches concentrations normally found only in females and in some species nanomolar concentrations of estrone sulfate are found in semen. Aromatase, which converts androgens to estrogens, is expressed in Leydig cells, seminiferous epithelium, and other male organs. Early studies showed E2 binding in numerous male tissues, and ESR1 and ESR2 each show unique distributions and actions in males. Exogenous estrogen treatment produced male reproductive pathologies in laboratory animals and men, especially during development, and studies with transgenic mice with compromised estrogen signaling demonstrated an E2 role in normal male physiology. Efferent ductules and epididymal functions are dependent on estrogen signaling through ESR1, whose loss impaired ion transport and water reabsorption, resulting in abnormal sperm. Loss of ESR1 or aromatase also produces effects on nonreproductive targets such as brain, adipose, skeletal muscle, bone, cardiovascular, and immune tissues. Expression of GPER is extensive in male tracts, suggesting a possible role for E2 signaling through this receptor in male reproduction. Recent evidence also indicates that membrane ESR1 has critical roles in male reproduction. Thus estrogens are important physiological regulators in males, and future studies may reveal additional roles for estrogen signaling in various target tissues.
Copyright © 2017 the American Physiological Society.
Figures



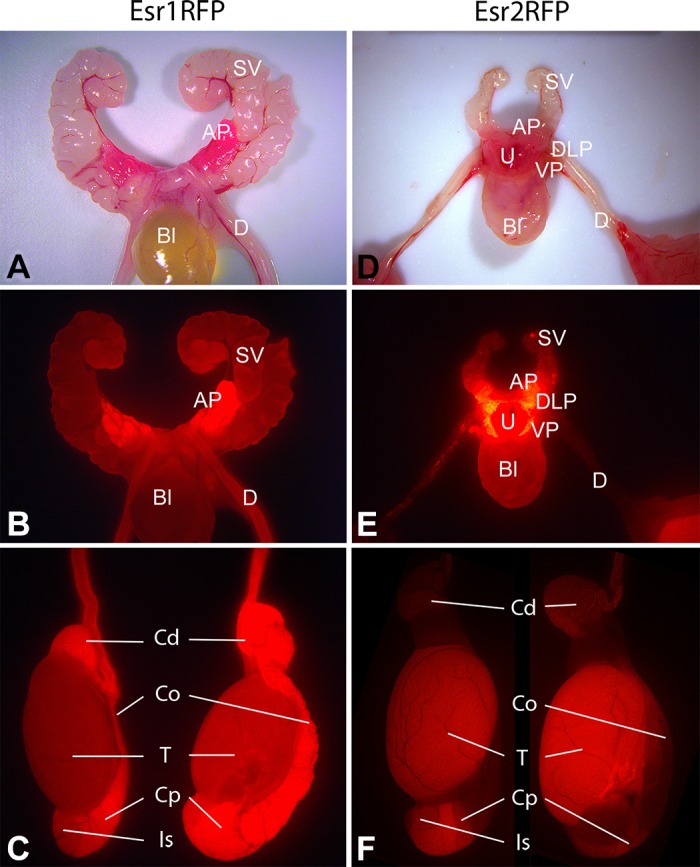


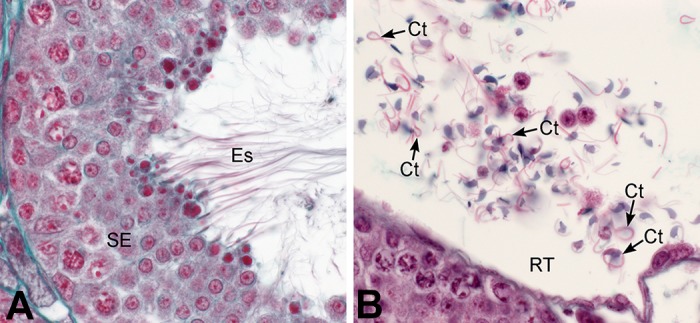


References
-
- Adlanmerini M, Solinhac R, Abot A, Fabre A, Raymond-Letron I, Guihot AL, Boudou F, Sautier L, Vessières E, Kim SH, Lière P, Fontaine C, Krust A, Chambon P, Katzenellenbogen JA, Gourdy P, Shaul PW, Henrion D, Arnal JF, Lenfant F. Mutation of the palmitoylation site of estrogen receptor α in vivo reveals tissue-specific roles for membrane versus nuclear actions. Proc Natl Acad Sci USA 111: E283–E290, 2014. doi: 10.1073/pnas.1322057111. - DOI - PMC - PubMed
-
- Agarwal VR, Takayama K, Van Wyk JJ, Sasano H, Simpson ER, Bulun SE. Molecular basis of severe gynecomastia associated with aromatase expression in a fibrolamellar hepatocellular carcinoma. J Clin Endocrinol Metab 83: 1797–1800, 1998. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous