

Independent evolution of baleen whale gigantism linked to Plio-Pleistocene ocean dynamics
- PMID: 28539520
- PMCID: PMC5454272
- DOI: 10.1098/rspb.2017.0546
Independent evolution of baleen whale gigantism linked to Plio-Pleistocene ocean dynamics
Abstract
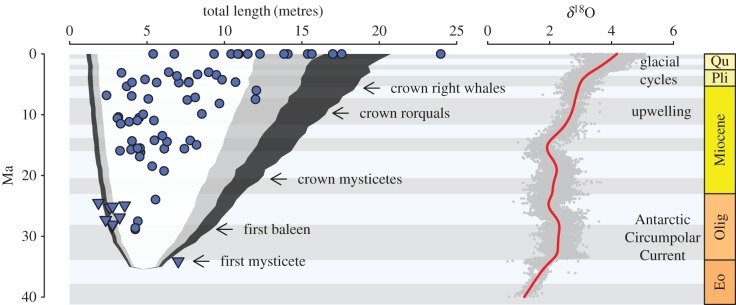
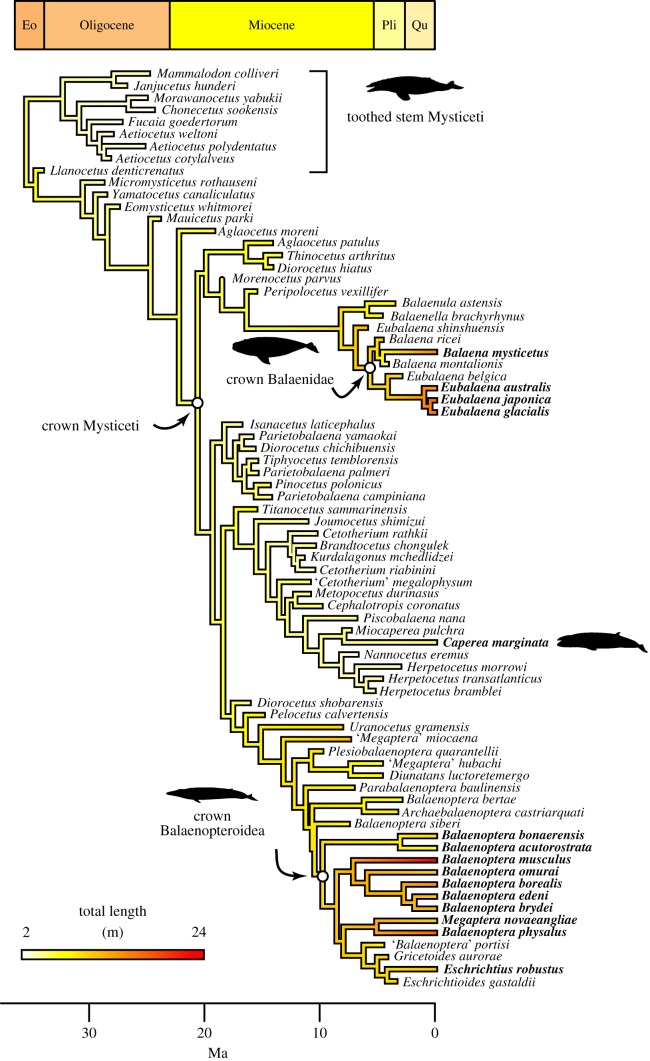
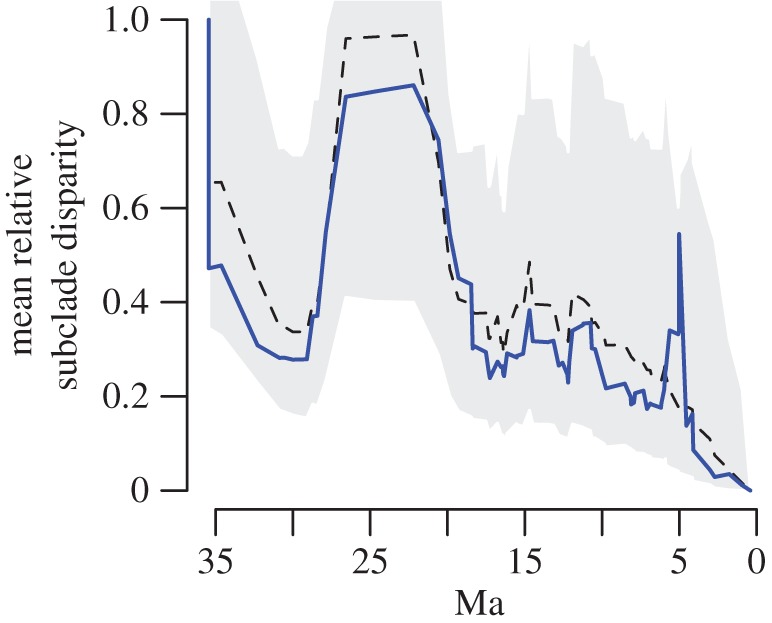
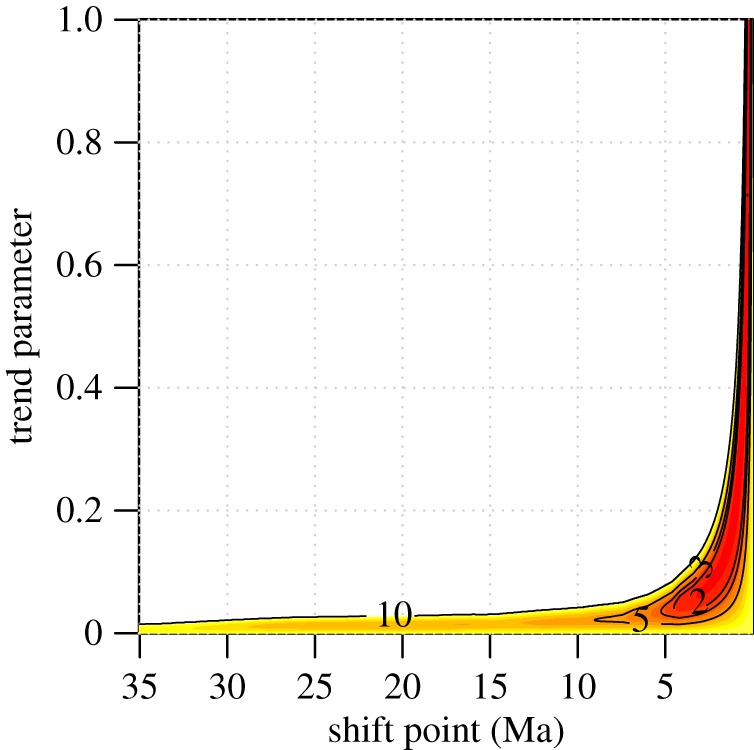
Vertebrates have evolved to gigantic sizes repeatedly over the past 250 Myr, reaching their extreme in today's baleen whales (Mysticeti). Hypotheses for the evolution of exceptionally large size in mysticetes range from niche partitioning to predator avoidance, but there has been no quantitative examination of body size evolutionary dynamics in this clade and it remains unclear when, why or how gigantism evolved. By fitting phylogenetic macroevolutionary models to a dataset consisting of living and extinct species, we show that mysticetes underwent a clade-wide shift in their mode of body size evolution during the Plio-Pleistocene. This transition, from Brownian motion-like dynamics to a trended random walk towards larger size, is temporally linked to the onset of seasonally intensified upwelling along coastal ecosystems. High prey densities resulting from wind-driven upwelling, rather than abundant resources alone, are the primary determinant of efficient foraging in extant mysticetes and Late Pliocene changes in ocean dynamics may have provided an ecological pathway to gigantism in multiple independent lineages.
Keywords: body size; fossil; macroevolution; phylogeny; tempo and mode; trend.
© 2017 The Author(s).
Conflict of interest statement
We declare we have no competing interests.
Figures
References
-
- Smith FA. et al. 2016. Body size evolution across the Geozoic. Annu. Rev. Earth Planet. Sci. 44, 523–553. ( 10.1146/annurev-earth-060115-012147) - DOI
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous