

The role of retinol dehydrogenase 10 in the cone visual cycle
- PMID: 28539612
- PMCID: PMC5443843
- DOI: 10.1038/s41598-017-02549-8
The role of retinol dehydrogenase 10 in the cone visual cycle
Abstract
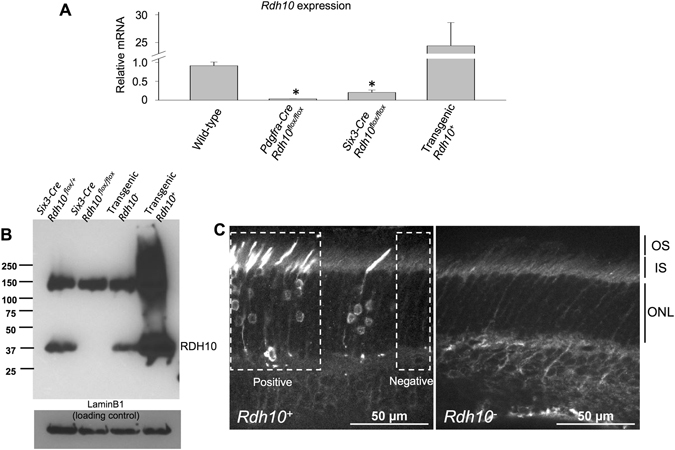
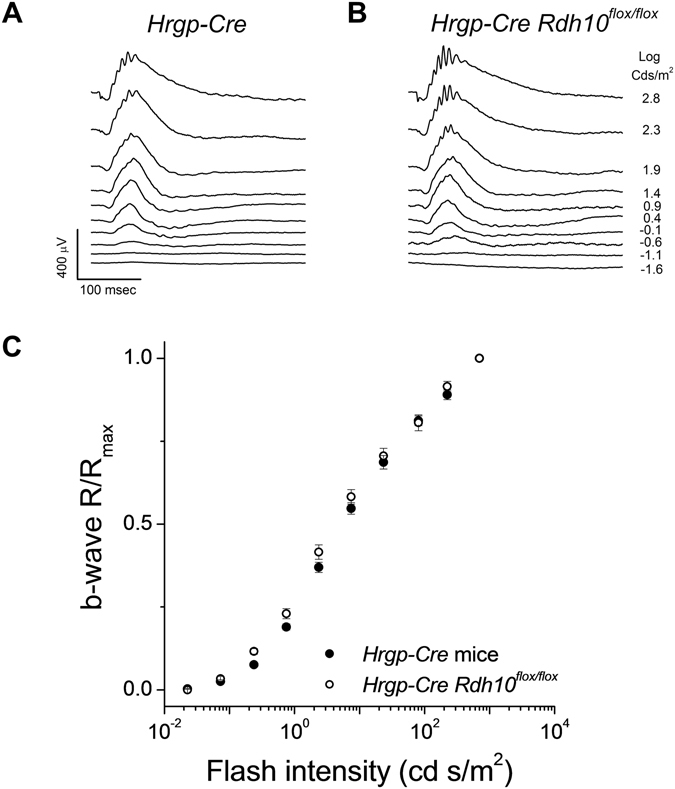
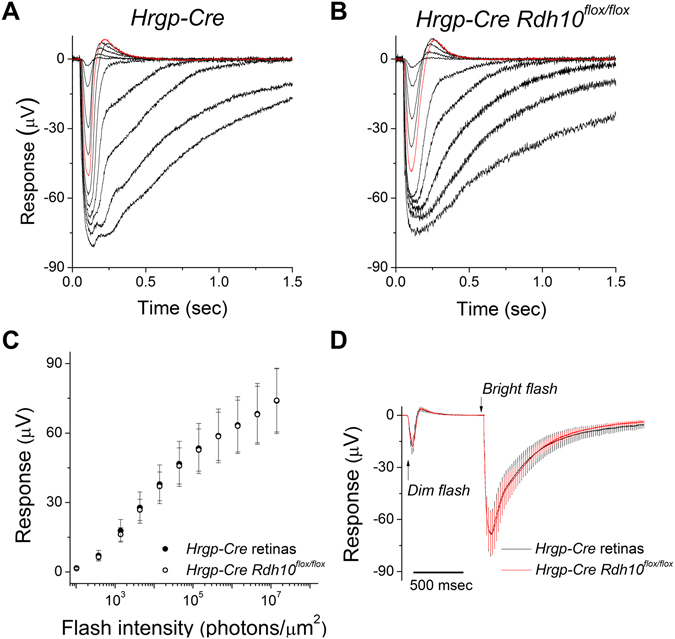
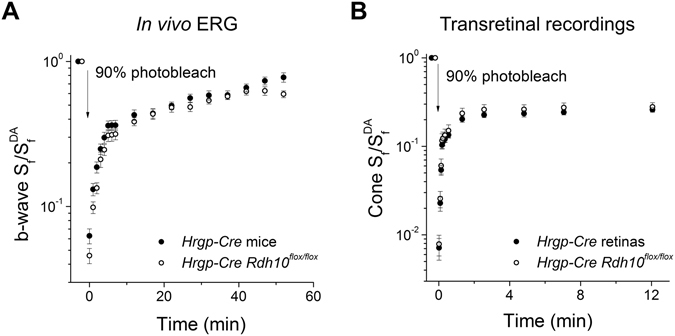
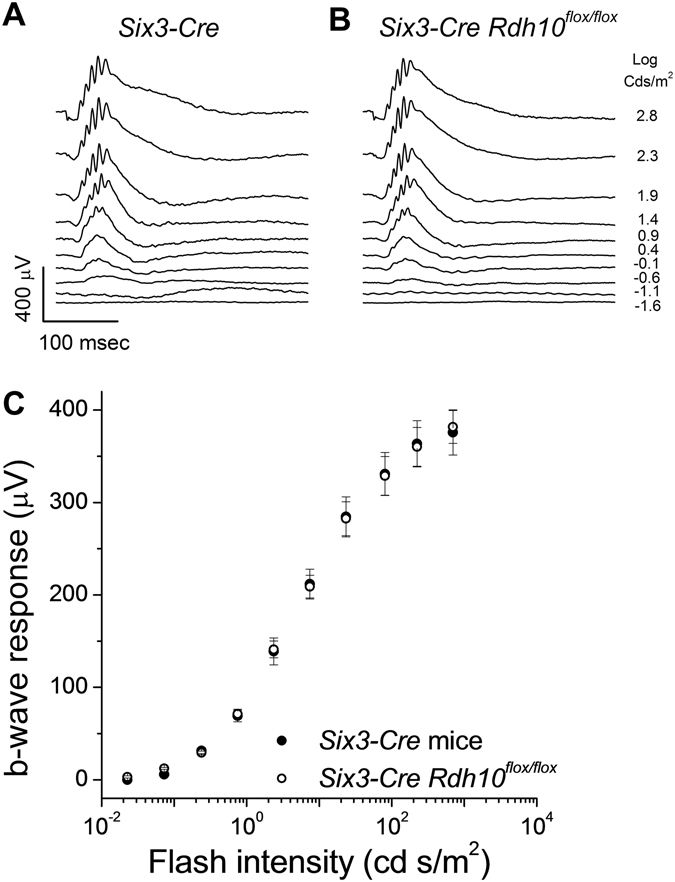
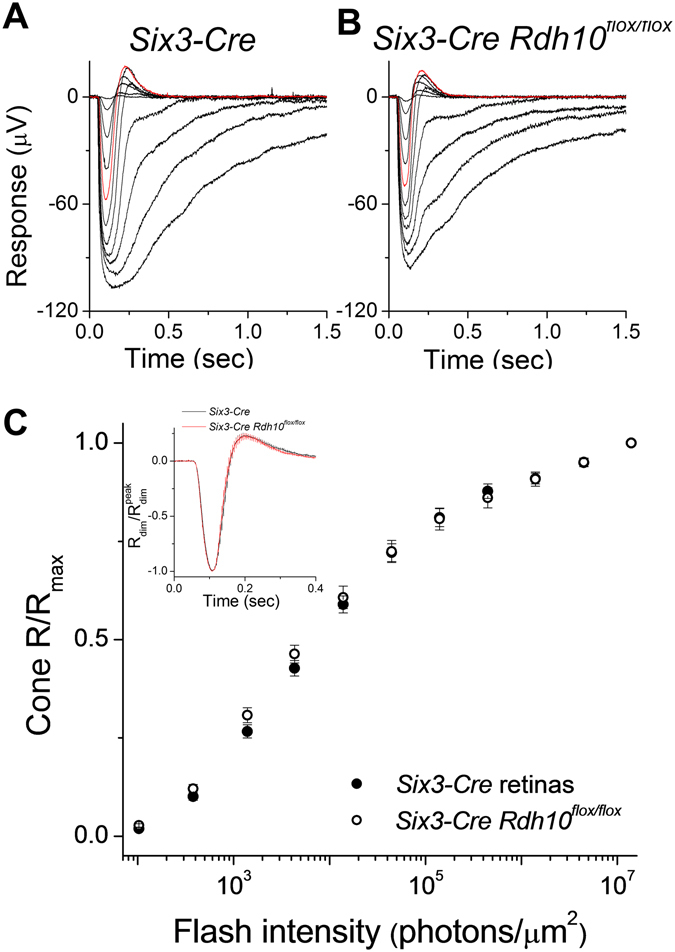
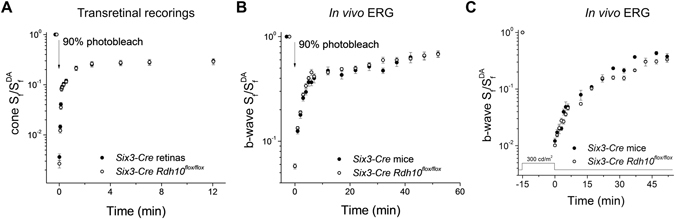
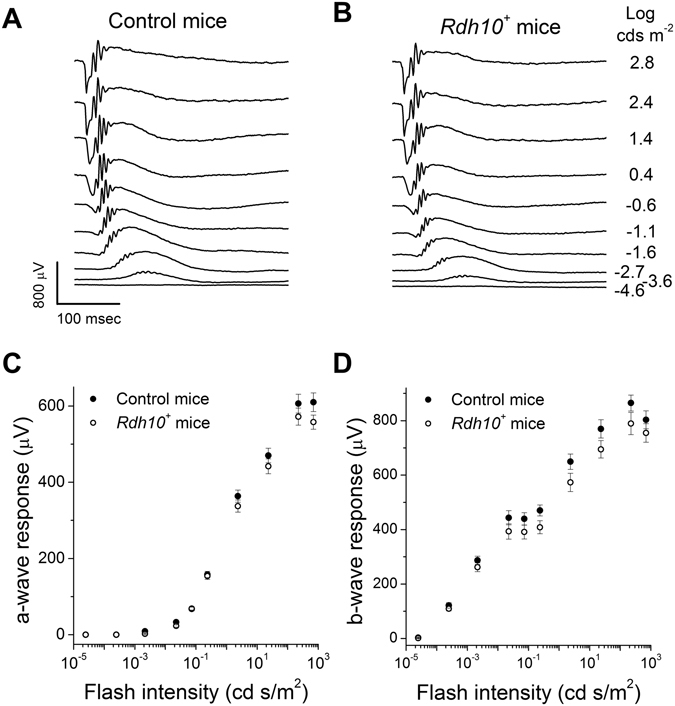
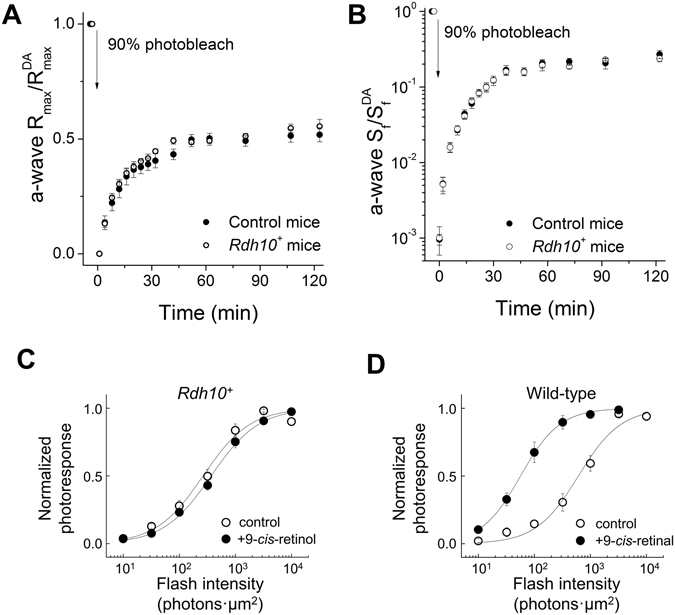
Pigment regeneration is critical for the function of cone photoreceptors in bright and rapidly-changing light conditions. This process is facilitated by the recently-characterized retina visual cycle, in which Müller cells recycle spent all-trans-retinol visual chromophore back to 11-cis-retinol. This 11-cis-retinol is oxidized selectively in cones to the 11-cis-retinal used for pigment regeneration. However, the enzyme responsible for the oxidation of 11-cis-retinol remains unknown. Here, we sought to determine whether retinol dehydrogenase 10 (RDH10), upregulated in rod/cone hybrid retinas and expressed abundantly in Müller cells, is the enzyme that drives this reaction. We created mice lacking RDH10 either in cone photoreceptors, Müller cells, or the entire retina. In vivo electroretinography and transretinal recordings revealed normal cone photoresponses in all RDH10-deficient mouse lines. Notably, their cone-driven dark adaptation both in vivo and in isolated retina was unaffected, indicating that RDH10 is not required for the function of the retina visual cycle. We also generated transgenic mice expressing RDH10 ectopically in rod cells. However, rod dark adaptation was unaffected by the expression of RDH10 and transgenic rods were unable to use cis-retinol for pigment regeneration. We conclude that RDH10 is not the dominant retina 11-cis-RDH, leaving its primary function in the retina unknown.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
Similar articles
-
Conditional deletion of Des1 in the mouse retina does not impair the visual cycle in cones.FASEB J. 2019 Apr;33(4):5782-5792. doi: 10.1096/fj.201802493R. Epub 2019 Jan 15. FASEB J. 2019. PMID: 30645148 Free PMC article.
-
cis Retinol oxidation regulates photoreceptor access to the retina visual cycle and cone pigment regeneration.J Physiol. 2016 Nov 15;594(22):6753-6765. doi: 10.1113/JP272831. Epub 2016 Aug 2. J Physiol. 2016. PMID: 27385534 Free PMC article.
-
Retinol dehydrogenase 8 and ATP-binding cassette transporter 4 modulate dark adaptation of M-cones in mammalian retina.J Physiol. 2015 Nov 15;593(22):4923-41. doi: 10.1113/JP271285. Epub 2015 Oct 18. J Physiol. 2015. PMID: 26350353 Free PMC article.
-
The visual cycle of the cone photoreceptors of the retina.Nutr Rev. 2004 Jul;62(7 Pt 1):283-6. doi: 10.1111/j.1753-4887.2004.tb00053.x. Nutr Rev. 2004. PMID: 15384919 Review.
-
Retinol Dehydrogenases Regulate Vitamin A Metabolism for Visual Function.Nutrients. 2016 Nov 22;8(11):746. doi: 10.3390/nu8110746. Nutrients. 2016. PMID: 27879662 Free PMC article. Review.
Cited by
-
Conditional deletion of Des1 in the mouse retina does not impair the visual cycle in cones.FASEB J. 2019 Apr;33(4):5782-5792. doi: 10.1096/fj.201802493R. Epub 2019 Jan 15. FASEB J. 2019. PMID: 30645148 Free PMC article.
-
Shedding new light on the generation of the visual chromophore.Proc Natl Acad Sci U S A. 2020 Aug 18;117(33):19629-19638. doi: 10.1073/pnas.2008211117. Epub 2020 Aug 5. Proc Natl Acad Sci U S A. 2020. PMID: 32759209 Free PMC article. Review.
-
Mislocalization of cone nuclei impairs cone function in mice.FASEB J. 2020 Aug;34(8):10242-10249. doi: 10.1096/fj.202000568R. Epub 2020 Jun 15. FASEB J. 2020. PMID: 32539195 Free PMC article.
-
Retinoid dynamics in vision: from visual cycle biology to retina disease treatments.Pharmacol Ther. 2025 Sep;273:108902. doi: 10.1016/j.pharmthera.2025.108902. Epub 2025 Jun 21. Pharmacol Ther. 2025. PMID: 40550364 Free PMC article. Review.
-
AAV cis-regulatory sequences are correlated with ocular toxicity.Proc Natl Acad Sci U S A. 2019 Mar 19;116(12):5785-5794. doi: 10.1073/pnas.1821000116. Epub 2019 Mar 4. Proc Natl Acad Sci U S A. 2019. PMID: 30833387 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
