

Modulation of liver regeneration via myeloid PTEN deficiency
- PMID: 28542148
- PMCID: PMC5520744
- DOI: 10.1038/cddis.2017.47
Modulation of liver regeneration via myeloid PTEN deficiency
Abstract
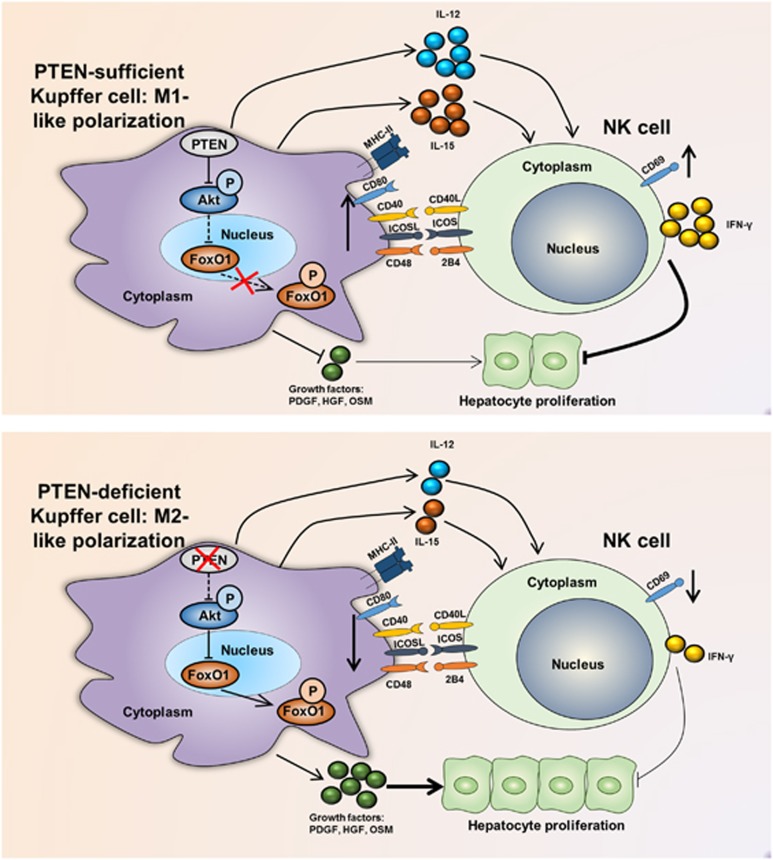
Molecular mechanisms that modulate liver regeneration are of critical importance for a number of hepatic disorders. Kupffer cells and natural killer (NK) cells are two cell subsets indispensable for liver regeneration. We have focused on these two populations and, in particular, the interplay between them. Importantly, we demonstrate that deletion of the myeloid phosphatase and tensin homolog on chromosome 10 (PTEN) leading to an M2-like polarization of Kupffer cells, which results in decreased activation of NK cells. In addition, PTEN-deficient Kupffer cells secrete additional factors that facilitate the proliferation of hepatocytes. In conclusion, PTEN is critical for inhibiting M2-like polarization of Kupffer cells after partial hepatectomy, resulting in NK cell activation and thus the inhibition of liver regeneration. Furthermore, PTEN reduces growth factor secretion by Kupffer cells. Our results suggest that targeting PTEN on Kupffer cells may be useful in altering liver regeneration in patients undergoing liver resection.
Conflict of interest statement
The authors declare no conflict of interest.
Figures






Similar articles
-
PTEN Down-Regulation Promotes β-Oxidation to Fuel Hypertrophic Liver Growth After Hepatectomy in Mice.Hepatology. 2017 Sep;66(3):908-921. doi: 10.1002/hep.29226. Epub 2017 Jul 20. Hepatology. 2017. PMID: 28437835
-
MicroRNA-21 Contributes to Liver Regeneration by Targeting PTEN.Med Sci Monit. 2016 Jan 8;22:83-91. doi: 10.12659/MSM.896157. Med Sci Monit. 2016. PMID: 26744142 Free PMC article.
-
The role of PTEN in regulation of hepatic macrophages activation and function in progression and reversal of liver fibrosis.Toxicol Appl Pharmacol. 2017 Feb 15;317:51-62. doi: 10.1016/j.taap.2017.01.005. Epub 2017 Jan 14. Toxicol Appl Pharmacol. 2017. PMID: 28095306
-
Cooperation of liver cells in health and disease.Adv Anat Embryol Cell Biol. 2001;161:III-XIII, 1-151. doi: 10.1007/978-3-642-56553-3. Adv Anat Embryol Cell Biol. 2001. PMID: 11729749 Review.
-
Kupffer cells, the limelight in the liver regeneration.Int Immunopharmacol. 2025 Jan 27;146:113808. doi: 10.1016/j.intimp.2024.113808. Epub 2024 Dec 13. Int Immunopharmacol. 2025. PMID: 39673997 Review.
Cited by
-
Cell Sources and Influencing Factors of Liver Regeneration: A Review.Med Sci Monit. 2020 Dec 14;26:e929129. doi: 10.12659/MSM.929129. Med Sci Monit. 2020. PMID: 33311428 Free PMC article. Review.
-
Global research trends in liver regeneration and immunomodulation: A perspective from bibliometric analysis.Hum Vaccin Immunother. 2025 Dec;21(1):2524249. doi: 10.1080/21645515.2025.2524249. Epub 2025 Jul 1. Hum Vaccin Immunother. 2025. PMID: 40591402 Free PMC article.
-
PTEN-mediated AKT/β-catenin signaling enhances the proliferation and expansion of Lgr5+ hepatocytes.Int J Biol Sci. 2021 Feb 17;17(3):861-868. doi: 10.7150/ijbs.56091. eCollection 2021. Int J Biol Sci. 2021. PMID: 33767594 Free PMC article.
-
Myeloid PTEN promotes chemotherapy-induced NLRP3-inflammasome activation and antitumour immunity.Nat Cell Biol. 2020 Jun;22(6):716-727. doi: 10.1038/s41556-020-0510-3. Epub 2020 May 4. Nat Cell Biol. 2020. PMID: 32367047
-
Molecular mechanisms in liver repair and regeneration: from physiology to therapeutics.Signal Transduct Target Ther. 2025 Feb 8;10(1):63. doi: 10.1038/s41392-024-02104-8. Signal Transduct Target Ther. 2025. PMID: 39920130 Free PMC article. Review.
References
-
- Selzner N, Selzner M, Odermatt B, Tian YH, Van Rooijen N, Clavien PA. ICAM-1 triggers liver regeneration through leukocyte recruitment and Kupffer cell-dependent release of TNF-alpha/IL-6 in mice. Gastroenterology 2003; 124: 692–700. - PubMed
-
- Fausto N, Campbell JS, Riehle KJ. Liver regeneration. Hepatology 2006; 43: S45–S53. - PubMed
-
- Elsegood CL, Chan CW, Degli-Esposti MA, Wikstrom ME, Domenichini A, Lazarus K et al. Kupffer cell-monocyte communication is essential for initiating murine liver progenitor cell-mediated liver regeneration. Hepatology 2015; 62: 1272–1284. - PubMed
-
- Sun R, Gao B. Negative regulation of liver regeneration by innate immunity (natural killer cells/interferon-gamma). Gastroenterology 2004; 127: 1525–1539. - PubMed
-
- Bi JC, Zheng XD, Chen YY, Wei HM, Sun R, Tian ZG. TIGIT safeguards liver regeneration through regulating natural killer cell-hepatocyte crosstalk. Hepatology 2014; 60: 1389–1398. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
