

A human monocytic NF-κB fluorescent reporter cell line for detection of microbial contaminants in biological samples
- PMID: 28542462
- PMCID: PMC5443541
- DOI: 10.1371/journal.pone.0178220
A human monocytic NF-κB fluorescent reporter cell line for detection of microbial contaminants in biological samples
Abstract
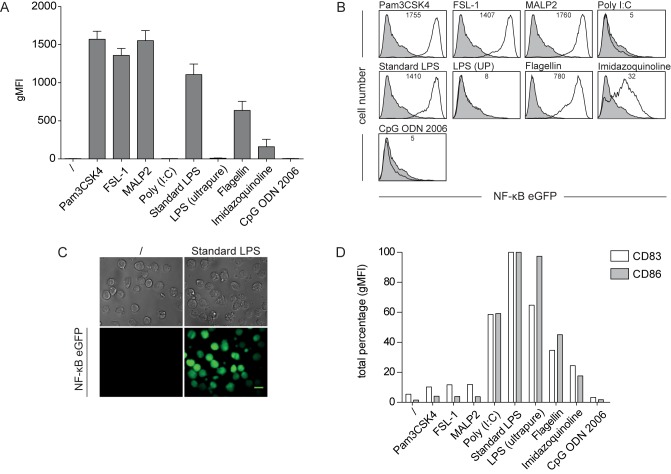
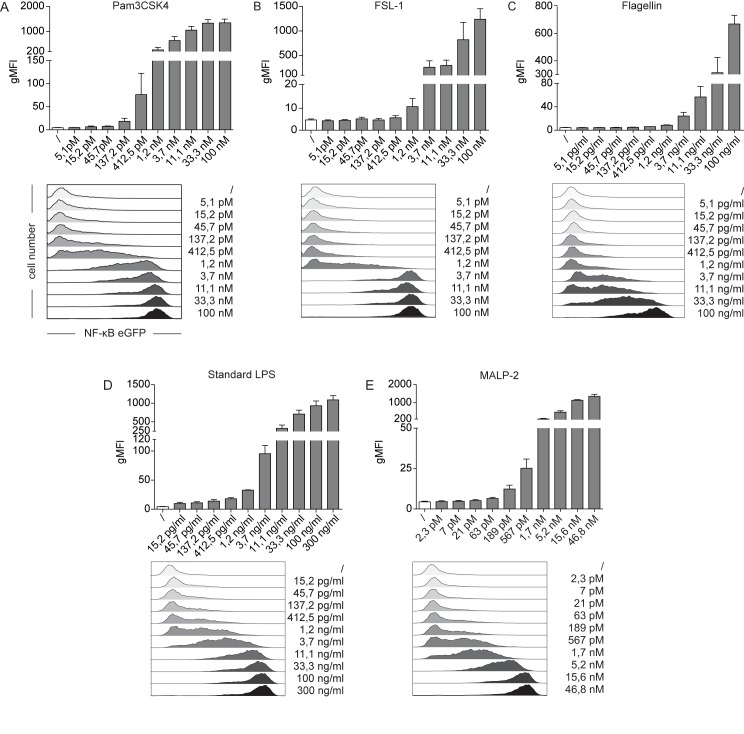
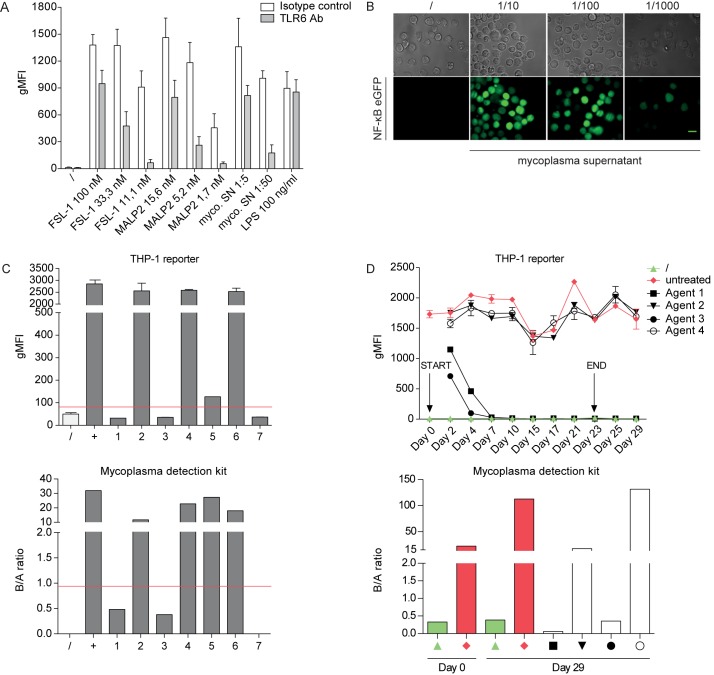
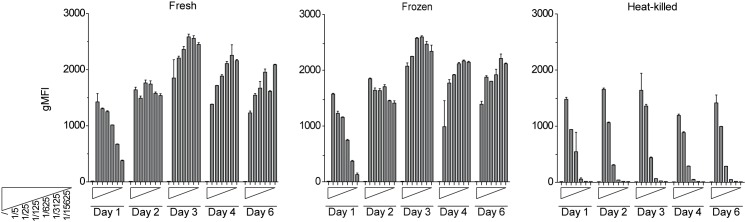
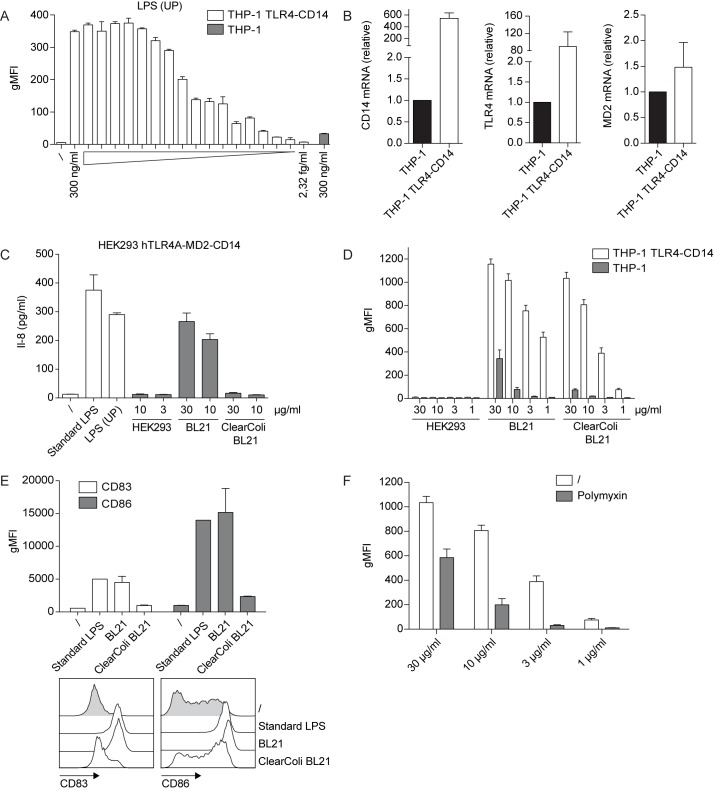
Sensing of pathogens by innate immune cells is essential for the initiation of appropriate immune responses. Toll-like receptors (TLRs), which are highly sensitive for various structurally and evolutionary conserved molecules derived from microbes have a prominent role in this process. TLR engagement results in the activation of the transcription factor NF-κB, which induces the expression of cytokines and other inflammatory mediators. The exquisite sensitivity of TLR signalling can be exploited for the detection of bacteria and microbial contaminants in tissue cultures and in protein preparations. Here we describe a cellular reporter system for the detection of TLR ligands in biological samples. The well-characterized human monocytic THP-1 cell line was chosen as host for an NF-ᴋB-inducible enhanced green fluorescent protein reporter gene. We studied the sensitivity of the resultant reporter cells for a variety of microbial components and observed a strong reactivity towards TLR1/2 and TLR2/6 ligands. Mycoplasma lipoproteins are potent TLR2/6 agonists and we demonstrate that our reporter cells can be used as reliable and robust detection system for mycoplasma contaminations in cell cultures. In addition, a TLR4-sensitive subline of our reporters was engineered, and probed with recombinant proteins expressed in different host systems. Bacterially expressed but not mammalian expressed proteins induced strong reporter activity. We also tested proteins expressed in an E. coli strain engineered to lack TLR4 agonists. Such preparations also induced reporter activation in THP-1 cells highlighting the importance of testing recombinant protein preparations for microbial contaminations beyond endotoxins. Our results demonstrate the usefulness of monocytic reporter cells for high-throughput screening for microbial contaminations in diverse biological samples, including tissue culture supernatants and recombinant protein preparations. Fluorescent reporter assays can be measured on standard flow cytometers and in contrast to established detection methods, like luciferase-based systems or Limulus Amebocyte Lysate tests, they do not require costly reagents.
Conflict of interest statement
Figures
References
-
- Cooper JF, Levin J, Wagner HN Jr. Quantitative comparison of in vitro and in vivo methods for the detection of endotoxin. The Journal of laboratory and clinical medicine. 1971;78(1):138–48. - PubMed
-
- Drexler HG, Uphoff CC. Mycoplasma contamination of cell cultures: Incidence, sources, effects, detection, elimination, prevention. Cytotechnology. 2002;39(2):75–90. doi: 10.1023/A:1022913015916 - DOI - PMC - PubMed
-
- Lemaitre B, Nicolas E, Michaut L, Reichhart JM, Hoffmann JA. The dorsoventral regulatory gene cassette spatzle/Toll/cactus controls the potent antifungal response in Drosophila adults. Cell. 1996;86(6):973–83. - PubMed
-
- Gay NJ, Keith FJ. Drosophila Toll and IL-1 receptor. Nature. 1991;351(6325):355–6. doi: 10.1038/351355b0 - DOI - PubMed
-
- Medzhitov R, Preston-Hurlburt P, Janeway CA Jr. A human homologue of the Drosophila Toll protein signals activation of adaptive immunity. Nature. 1997;388(6640):394–7. doi: 10.1038/41131 - DOI - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
