

UGGT1 enhances enterovirus 71 pathogenicity by promoting viral RNA synthesis and viral replication
- PMID: 28545059
- PMCID: PMC5435352
- DOI: 10.1371/journal.ppat.1006375
UGGT1 enhances enterovirus 71 pathogenicity by promoting viral RNA synthesis and viral replication
Abstract
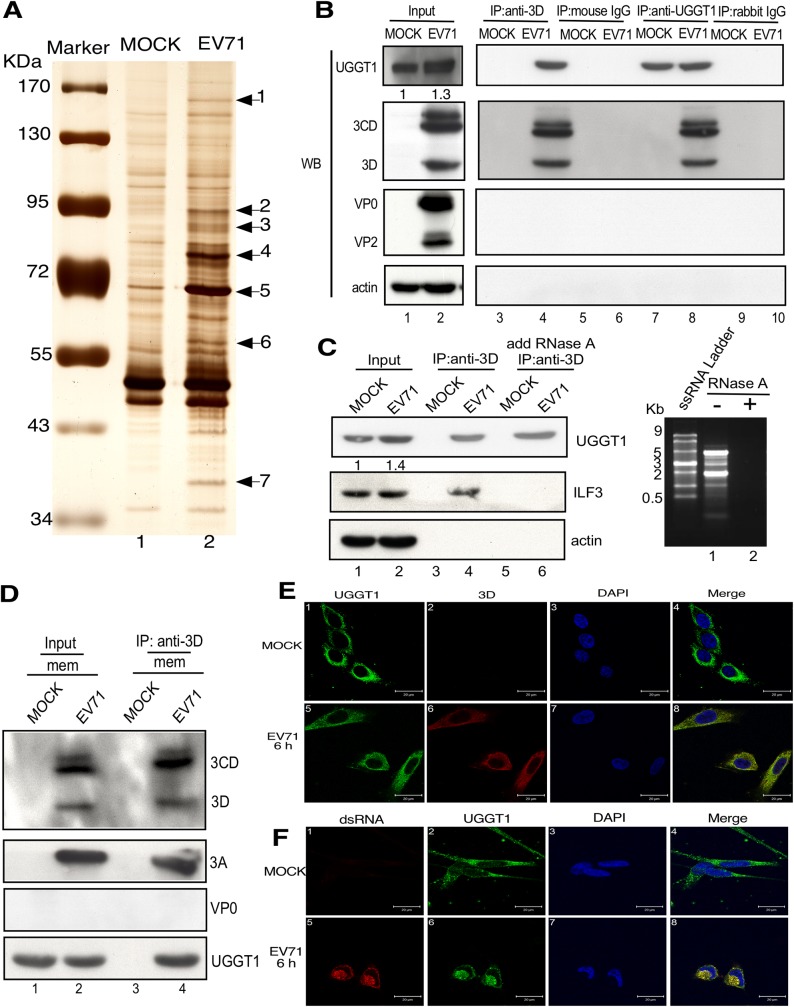
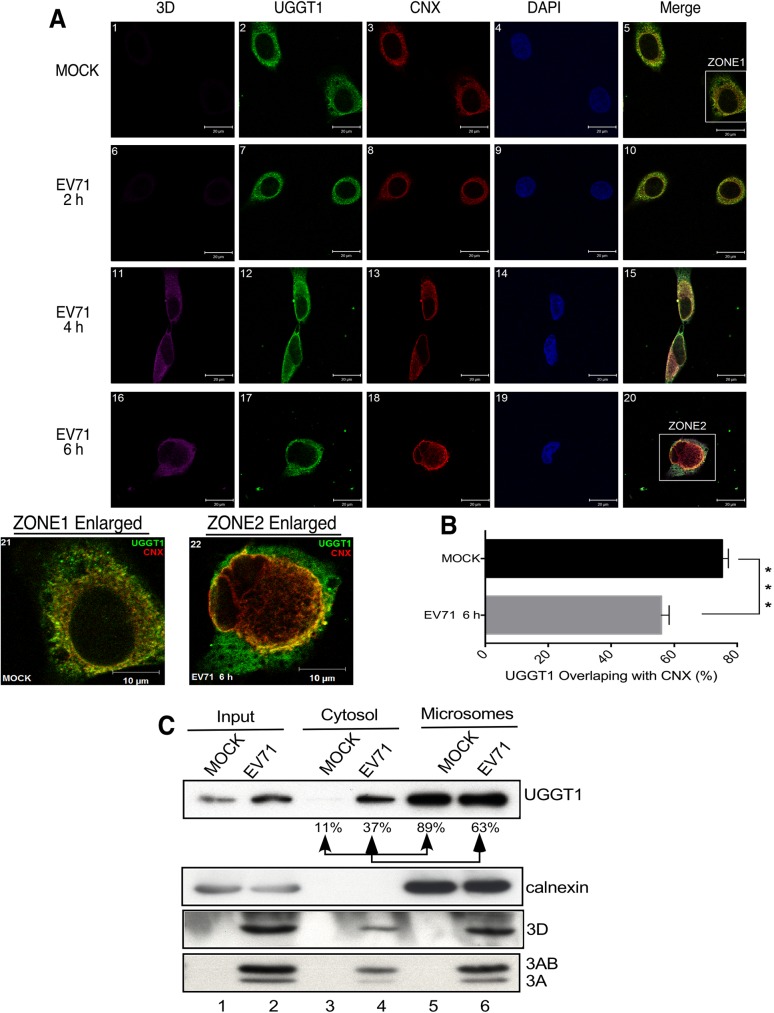
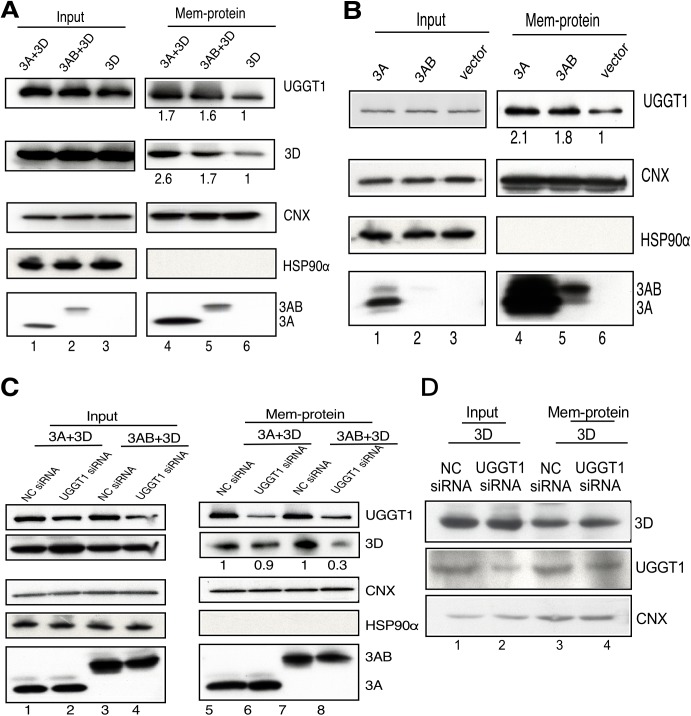
Positive-strand RNA virus infections can induce the stress-related unfolded protein response (UPR) in host cells. This study found that enterovirus A71 (EVA71) utilizes host UDP-glucose glycoprotein glucosyltransferase 1 (UGGT1), a key endoplasmic reticulum protein (ER) involved in UPR, to enhance viral replication and virulence. EVA71 forms replication complexes (RCs) on cellular membranes that contain a mix of host and viral proteins to facilitate viral replication, but the components and processes involved in the assembly and function of RCs are not fully understood. Using EVA71 as a model, this study found that host UGGT1 and viral 3D polymerase co-precipitate along with other factors on membranous replication complexes to enhance viral replication. Increased UGGT1 levels elevated viral growth rates, while viral pathogenicity was observed to be lower in heterozygous knockout mice (Uggt1 +/- mice). These findings provide important insight on the role of UPR and host UGGT1 in regulating RNA virus replication and pathogenicity.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures




Similar articles
-
Human Herpesvirus 8 Interleukin-6 Interacts with Calnexin Cycle Components and Promotes Protein Folding.J Virol. 2017 Oct 27;91(22):e00965-17. doi: 10.1128/JVI.00965-17. Print 2017 Nov 15. J Virol. 2017. PMID: 28878084 Free PMC article.
-
SUMO Modification Stabilizes Enterovirus 71 Polymerase 3D To Facilitate Viral Replication.J Virol. 2016 Nov 14;90(23):10472-10485. doi: 10.1128/JVI.01756-16. Print 2016 Dec 1. J Virol. 2016. PMID: 27630238 Free PMC article.
-
Secretory Carrier Membrane Protein 3 Interacts with 3A Viral Protein of Enterovirus and Participates in Viral Replication.Microbiol Spectr. 2021 Sep 3;9(1):e0047521. doi: 10.1128/Spectrum.00475-21. Epub 2021 Aug 11. Microbiol Spectr. 2021. PMID: 34378951 Free PMC article.
-
How does the stressed out ER find relief during virus infection?Curr Opin Virol. 2016 Apr;17:74-79. doi: 10.1016/j.coviro.2016.01.018. Epub 2016 Feb 9. Curr Opin Virol. 2016. PMID: 26871502 Review.
-
[Molecular Mechanism of Action of hnRNP K and RTN3 in the Replication of Enterovirus 71].Bing Du Xue Bao. 2015 Mar;31(2):197-200. Bing Du Xue Bao. 2015. PMID: 26164948 Review. Chinese.
Cited by
-
Return of the Neurotropic Enteroviruses: Co-Opting Cellular Pathways for Infection.Viruses. 2021 Jan 22;13(2):166. doi: 10.3390/v13020166. Viruses. 2021. PMID: 33499355 Free PMC article. Review.
-
Rescue of secretion of rare-disease-associated misfolded mutant glycoproteins in UGGT1 knock-out mammalian cells.Traffic. 2024 Jan;25(1):e12927. doi: 10.1111/tra.12927. Traffic. 2024. PMID: 38272446 Free PMC article.
-
Reflux of Endoplasmic Reticulum proteins to the cytosol inactivates tumor suppressors.EMBO Rep. 2021 May 5;22(5):e51412. doi: 10.15252/embr.202051412. Epub 2021 Mar 12. EMBO Rep. 2021. PMID: 33710763 Free PMC article.
-
SR-BI Interactome Analysis Reveals a Proviral Role for UGGT1 in Hepatitis C Virus Entry.Front Microbiol. 2019 Sep 6;10:2043. doi: 10.3389/fmicb.2019.02043. eCollection 2019. Front Microbiol. 2019. PMID: 31551978 Free PMC article.
-
The RNA-dependent RNA polymerase of enterovirus A71 associates with ribosomal proteins and positively regulates protein translation.RNA Biol. 2020 Apr;17(4):608-622. doi: 10.1080/15476286.2020.1722448. Epub 2020 Feb 6. RNA Biol. 2020. PMID: 32009553 Free PMC article.
References
-
- Belov GA, van Kuppeveld FJ (2012) (+)RNA viruses rewire cellular pathways to build replication organelles. Curr Opin Virol 2: 740–747. doi: 10.1016/j.coviro.2012.09.006 - DOI - PMC - PubMed
-
- Koonin EV, Dolja VV (1993) Evolution and taxonomy of positive-strand RNA viruses: implications of comparative analysis of amino acid sequences. Crit Rev Biochem Mol Biol 28: 375–430. doi: 10.3109/10409239309078440 - DOI - PubMed
-
- Koonin EV, Wolf YI, Nagasaki K, Dolja VV (2008) The Big Bang of picorna-like virus evolution antedates the radiation of eukaryotic supergroups. Nat Rev Microbiol 6: 925–939. doi: 10.1038/nrmicro2030 - DOI - PubMed
-
- McMinn PC (2002) An overview of the evolution of enterovirus 71 and its clinical and public health significance. FEMS Microbiol Rev 26: 91–107. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
