

Structures of the inactive and active states of RIP2 kinase inform on the mechanism of activation
- PMID: 28545134
- PMCID: PMC5436651
- DOI: 10.1371/journal.pone.0177161
Structures of the inactive and active states of RIP2 kinase inform on the mechanism of activation
Abstract
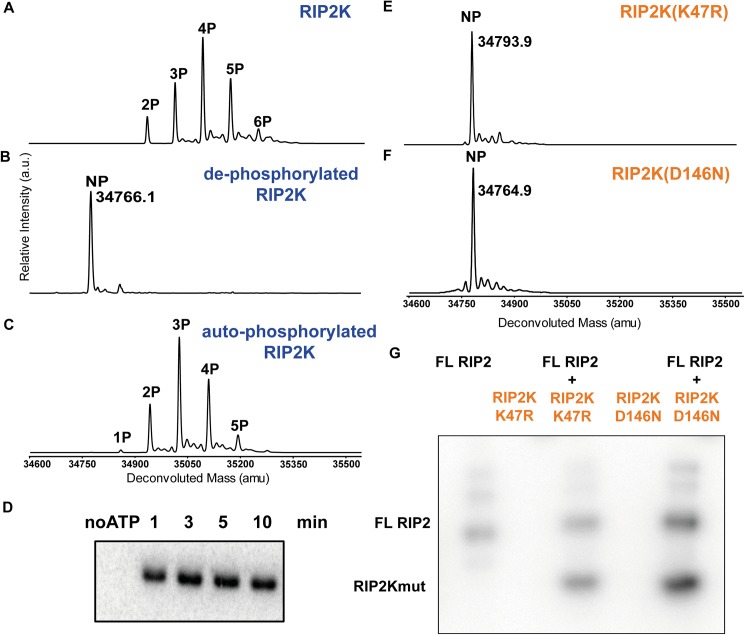
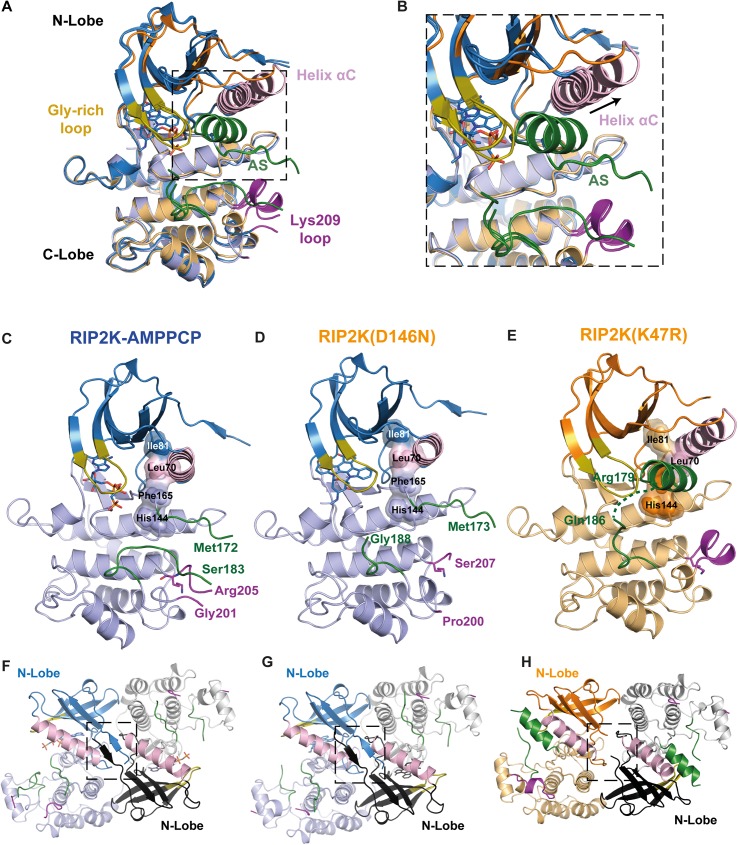
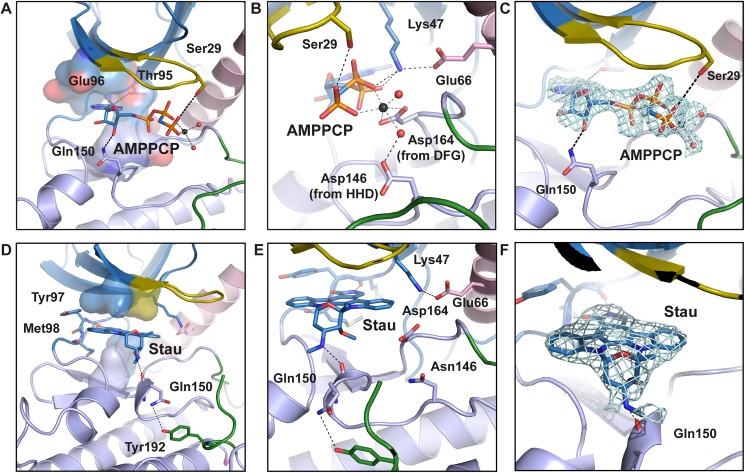
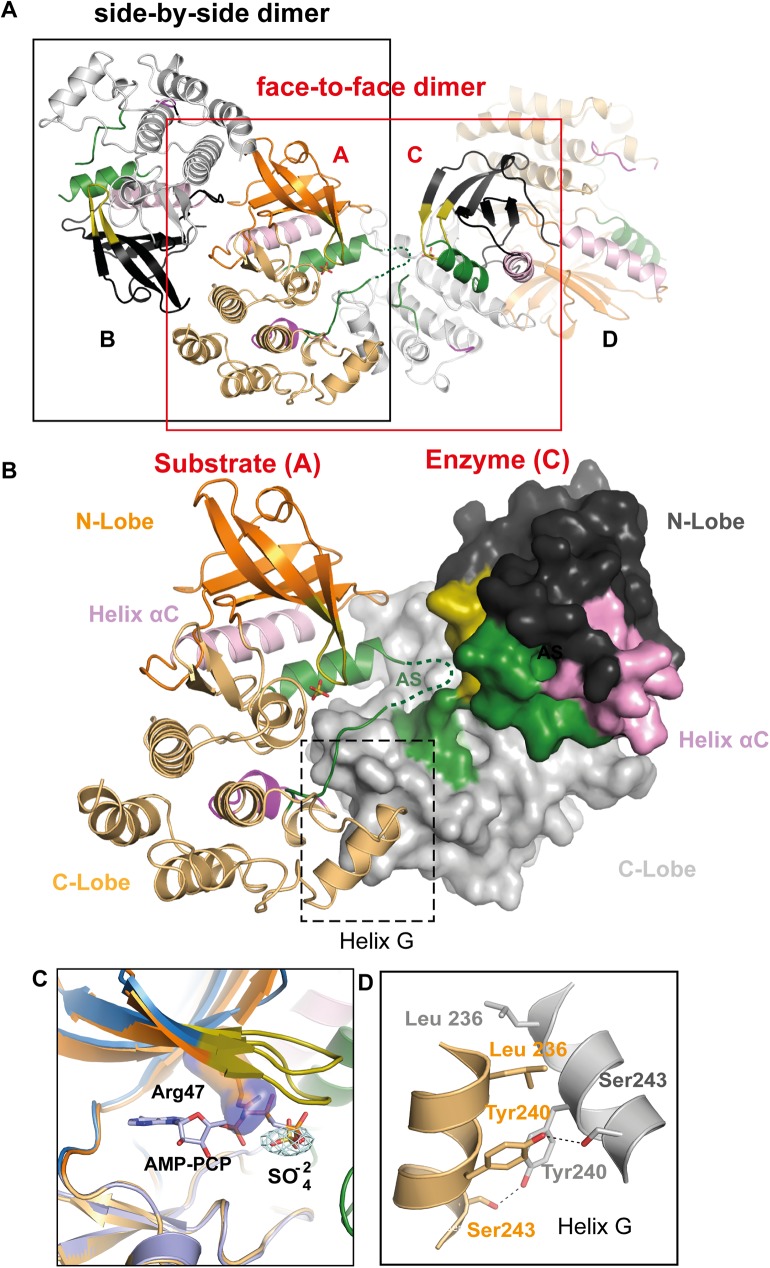
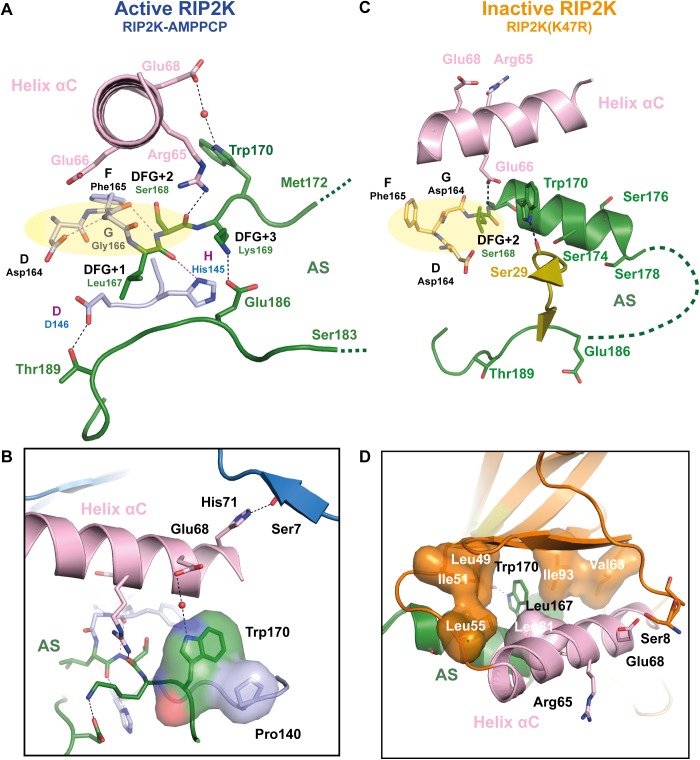
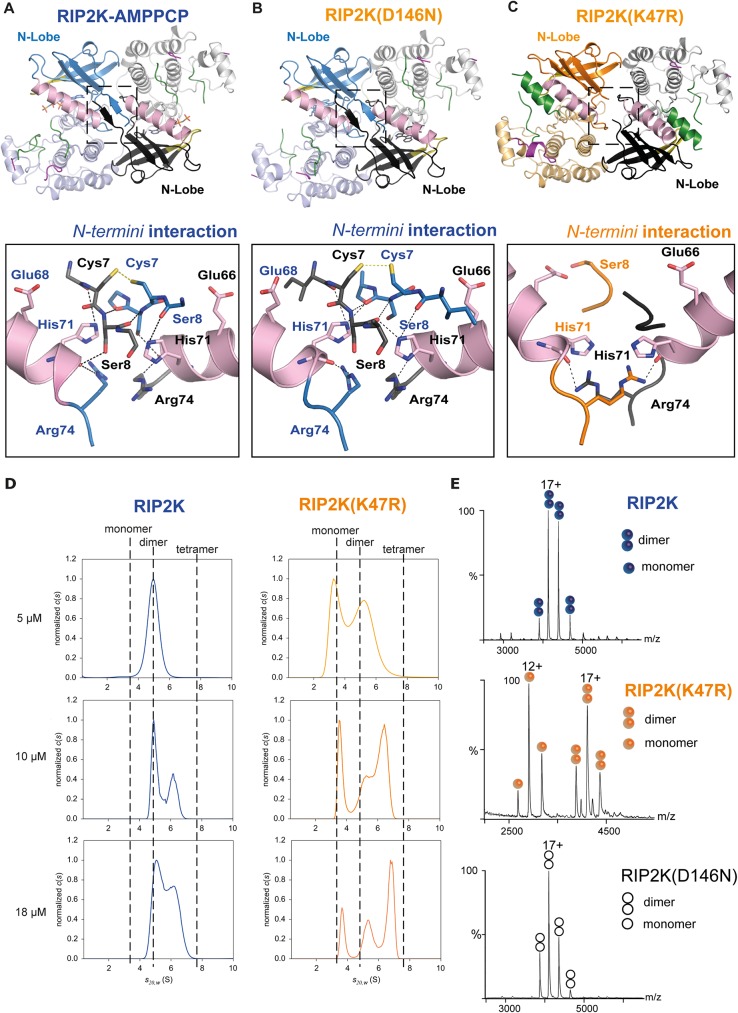
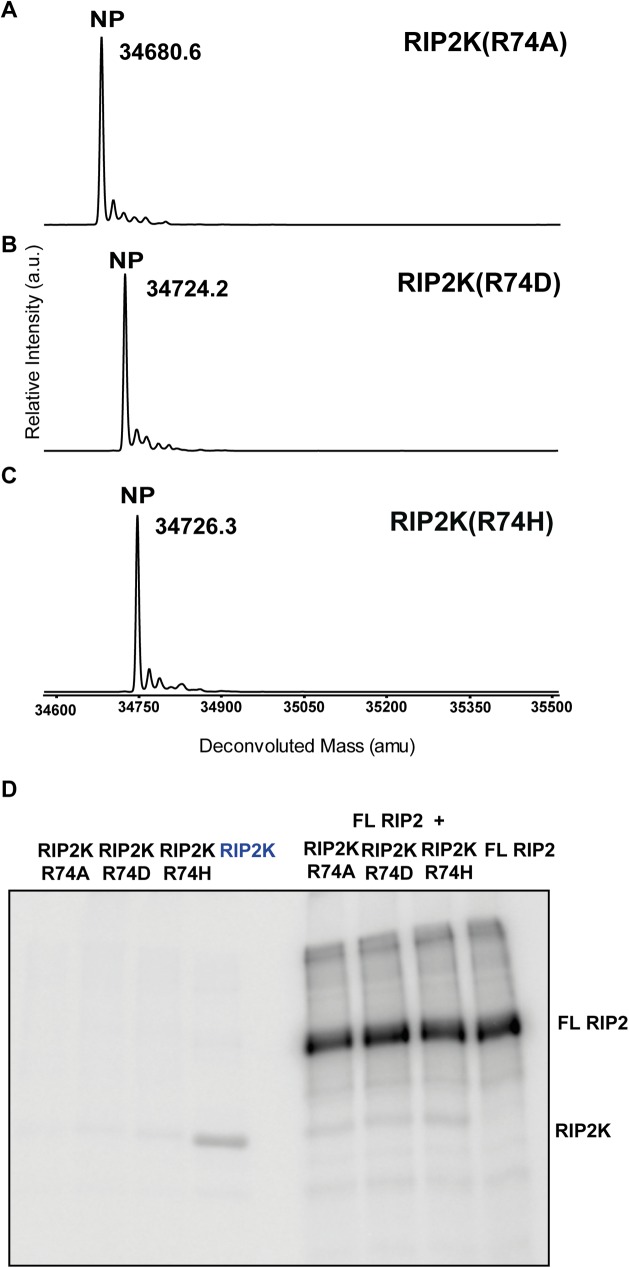
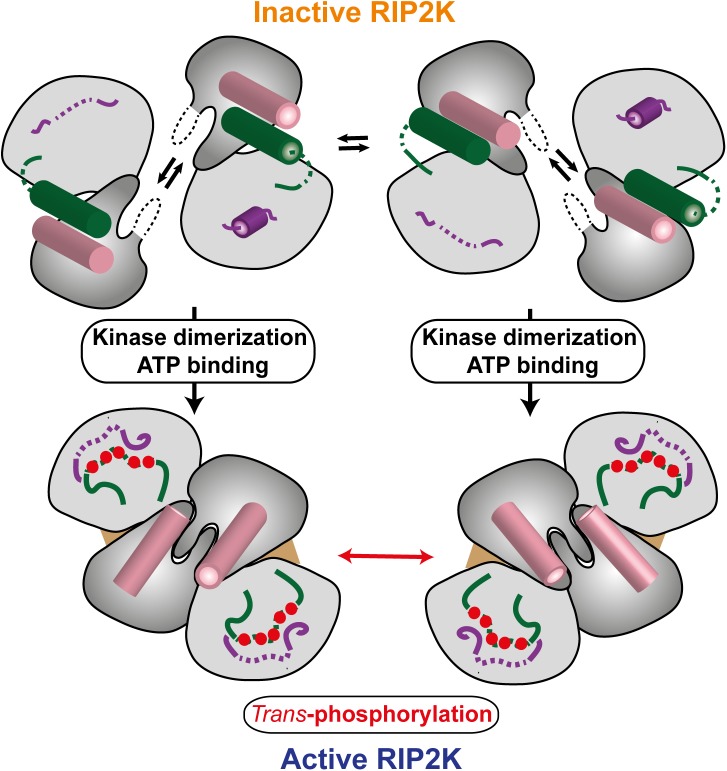
Innate immune receptors NOD1 and NOD2 are activated by bacterial peptidoglycans leading to recruitment of adaptor kinase RIP2, which, upon phosphorylation and ubiquitination, becomes a scaffold for downstream effectors. The kinase domain (RIP2K) is a pharmaceutical target for inflammatory diseases caused by aberrant NOD2-RIP2 signalling. Although structures of active RIP2K in complex with inhibitors have been reported, the mechanism of RIP2K activation remains to be elucidated. Here we analyse RIP2K activation by combining crystal structures of the active and inactive states with mass spectrometric characterization of their phosphorylation profiles. The active state has Helix αC inwardly displaced and the phosphorylated Activation Segment (AS) disordered, whilst in the inactive state Helix αC is outwardly displaced and packed against the helical, non-phosphorylated AS. Biophysical measurements show that the active state is a stable dimer whilst the inactive kinase is in a monomer-dimer equilibrium, consistent with the observed structural differences at the dimer interface. We conclude that RIP2 kinase auto-phosphorylation is intimately coupled to dimerization, similar to the case of BRAF. Our results will help drug design efforts targeting RIP2 as a potential treatment for NOD2-RIP2 related inflammatory diseases.
Conflict of interest statement
Figures
References
-
- Chen G, Shaw MH, Kim YG, Nunez G. NOD-like receptors: role in innate immunity and inflammatory disease. Annual review of pathology. 2009;4:365–98. doi: 10.1146/annurev.pathol.4.110807.092239 - DOI - PubMed
-
- Park JH, Kim YG, McDonald C, Kanneganti TD, Hasegawa M, Body-Malapel M, et al. RICK/RIP2 mediates innate immune responses induced through Nod1 and Nod2 but not TLRs. Journal of immunology. 2007;178(4):2380–6. - PubMed
-
- Grimes CL, Ariyananda Lde Z, Melnyk JE, O'Shea EK. The innate immune protein Nod2 binds directly to MDP, a bacterial cell wall fragment. Journal of the American Chemical Society. 2012;134(33):13535–7. PubMed Central PMCID: PMC3424850. doi: 10.1021/ja303883c - DOI - PMC - PubMed
-
- Girardin SE, Boneca IG, Viala J, Chamaillard M, Labigne A, Thomas G, et al. Nod2 is a general sensor of peptidoglycan through muramyl dipeptide (MDP) detection. The Journal of biological chemistry. 2003;278(11):8869–72. doi: 10.1074/jbc.C200651200 - DOI - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
