

The Golgi matrix protein giantin is required for normal cilia function in zebrafish
- PMID: 28546340
- PMCID: PMC5576078
- DOI: 10.1242/bio.025502
The Golgi matrix protein giantin is required for normal cilia function in zebrafish
Abstract
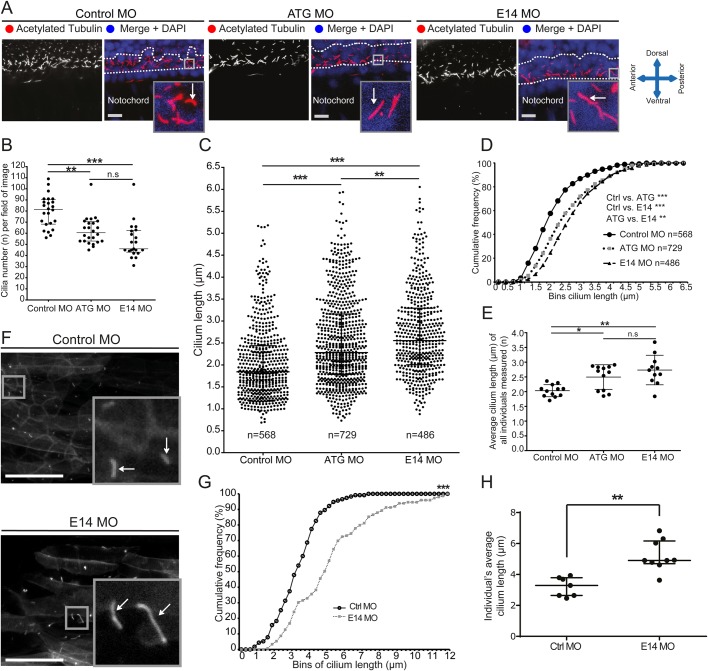
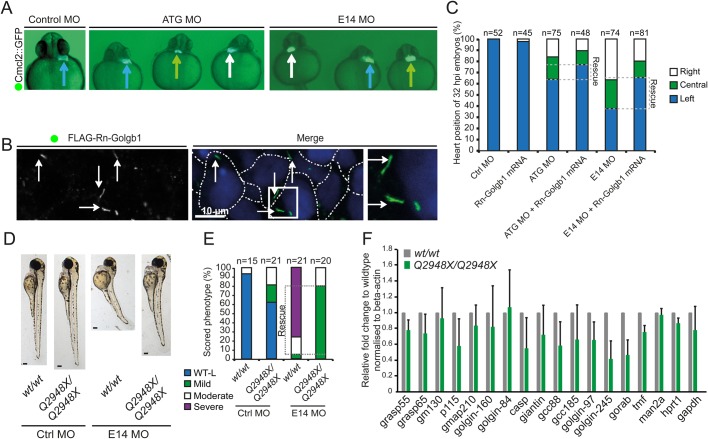
The Golgi is essential for glycosylation of newly synthesised proteins including almost all cell-surface and extracellular matrix proteoglycans. Giantin, encoded by the golgb1 gene, is a member of the golgin family of proteins that reside within the Golgi stack, but its function remains elusive. Loss of function of giantin in rats causes osteochondrodysplasia; knockout mice show milder defects, notably a cleft palate. In vitro, giantin has been implicated in Golgi organisation, biosynthetic trafficking, and ciliogenesis. Here we show that loss of function of giantin in zebrafish, using either morpholino or knockout techniques, causes defects in cilia function. Giantin morphants have fewer cilia in the neural tube and those remaining are longer. Mutants have the same number of cilia in the neural tube but these cilia are also elongated. Scanning electron microscopy shows that loss of giantin results in an accumulation of material at the ciliary tip, consistent with a loss of function of retrograde intraflagellar transport. Mutants show milder defects than morphants consistent with adaptation to loss of giantin.
Keywords: Cilia; Golgi; Golgi matrix; Zebrafish.
© 2017. Published by The Company of Biologists Ltd.
Conflict of interest statement
Competing interestsThe authors declare no competing or financial interests.
Figures



References
-
- Asante D., Maccarthy-Morrogh L., Townley A. K., Weiss M. A., Katayama K., Palmer K. J., Suzuki H., Westlake C. J. and Stephens D. J. (2013). A role for the Golgi matrix protein giantin in ciliogenesis through control of the localization of dynein-2. J. Cell Sci. 126, 5189-5197. 10.1242/jcs.131664 - DOI - PMC - PubMed
-
- Cermak T., Doyle E. L., Christian M., Wang L., Zhang Y., Schmidt C., Baller J. A., Somia N. V., Bogdanove A. J. and Voytas D. F. (2011). Efficient design and assembly of custom TALEN and other TAL effector-based constructs for DNA targeting. Nucleic Acids Res. 39, e82 10.1093/nar/gkr218 - DOI - PMC - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
